- 1. Nomenclature
- 2. Structure des acides nucléiques
- 3. Synthèse totale des acides nucléiques
- 4. Propriétés physico-chimiques
- 5. Dénaturation de l'ADN. Hybrides moléculaires
- 6. Rôle de l'acide désoxyribonucléique
- 7. Rôle des acides ribonucléiques
- 8. Biosynthèse des acides nucléiques
- 9. Lésions et réparations de l'ADN
- 10. Pathologies réparationnelles
- 11. Bibliographie
NUCLÉIQUES ACIDES
Article modifié le
Structure des acides nucléiques
Conformation et masse moléculaires des ARN
Les ARN messagers sont des molécules filamenteuses à chaîne simple, donc monocaténaires, plus ou moins longues et de stabilité souvent précaire.
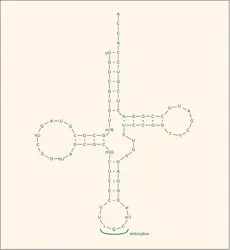
ARN de transfert : structure en feuille de trèfle
Encyclopædia Universalis France
Les ARN de transfert, dont on verra le rôle dans la dernière partie de cet article, sont constitués par l'enchaînement de quatre-vingts nucléotides environ, le tout ayant une masse moléculaire de l'ordre de 25 000. L'alanyl-ARN-t de levure représenté sur la figure 3 comporte soixante-dix-sept nucléotides et sa masse moléculaire est de 26 000.
Dès que cette séquence et celles d'autres ARN de transfert furent connues, on se rendit compte qu'il existait une possibilité d'appariement entre les bases (stabilisant la structure de la molécule) par l'intermédiaire de liaisons non covalentes entre A — U et G — C (mais à l'encontre de l'ADN, le couple A — U au lieu du couple A — T). Plusieurs modèles ont été proposés, mais il semble que le modèle « en feuille de trèfle » rende le mieux compte des résultats expérimentaux. Des modèles de structure à trois dimensions (structure tertiaire) ont été prévus.

D'autres ARN, à structure plus complexe que les ARN de transfert, c'est-à-dire par exemple l'ARN 5 S d'Escherichia coli, les ARN constitutifs des ribosomes, les ARN viraux, ont été également l'objet d'études assidues.
Les ARN à fonction régulatrice (micro-ARN, ARN interférant) ont été identifiés dès 1993 et en 2001 : leur utilisation par la biologie d’intervention (cf. biologie - Les pratiques interventionnelles) est en cours.
Conformation et masse moléculaires de l'ADN
L' ADN isolé à partir d'une source naturelle comme le thymus de veau se présente sous forme de fibres blanches et légères, comparables à première vue à de la cellulose. Sous forme de sel de sodium, l'ADN se dissout lentement dans l'eau pour former des solutions colloïdales extrêmement visqueuses.
Depuis le début de ce siècle, on sait que l'ADN contient quatre bases principales : adénine (A), cytosine (C), guanine (G) et thymine (T). Le dosage de ces bases par les anciennes méthodes fit croire à leur répartition équimoléculaire, d'où la célèbre théorie des tétranucléotides qui prévalut pendant longtemps et ne fut réfutée que vers les années 1948-1950 lorsque l'on sut doser avec plus de précision les purines et les pyrimidines. Ce fut l'application extensive de la chromatographie sur papier, notamment par E. Chargaff et ses élèves, qui permit de définir un véritable atlas de la constitution en nucléotides d'un grand nombre d'acides désoxyribonucléiques naturels.
La connaissance de la constitution en bases d'un ADN permet de déterminer la constitution globale de cet acide nucléique, puisque l'on sait par définition que chaque nucléotide comporte une base (à choisir parmi quatre), un pentose qui est toujours ici le désoxyribose et un groupement phosphoryle.
Une constatation fondamentale devait se faire jour progressivement : d'une part, l'égalité des proportions en adénine et thymine et en guanine et cytosine ; la constance du rapport (A + T)/(G + C) pour une même espèce animale, d'autre part. Cette constatation a pour corollaire immédiat que le rapport purines/pyrimidines est toujours égal à 1.
La nature de la liaison entre nucléotides fut longtemps controversée, mais, sur la foi d'études aux rayons X et d'études enzymatiques, on s'est arrêté finalement à la liaison 3′-5′ : comme on l'a déjà vu, le groupement phosphoryle établit un pont entre le groupement hydroxyle en position C-3′ d'un premier nucléotide et le groupement hydroxyle en position C-5′ d'un second nucléotide (fig. 2).
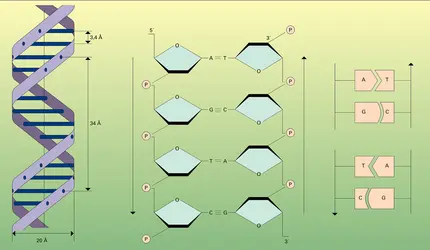
Structure bicaténaire de l'ADN
Encyclopædia Universalis France
Compte tenu des relations AT-GC, de la liaison 3′-5′-phosphodiester et des clichés obtenus aux rayons X, J. D. Watson et F. H. C. Crick proposèrent en 1952-1953 un schéma général de la conformation spatiale de l'ADN ; ce schéma, mettant en évidence le rôle génétique de cet acide nucléique, est à la base de la théorie de Watson et Crick ; il constitue l'un des plus grands événements scientifiques du xxe siècle (fig. 4 ; cf. macromolécules).
On doit se représenter la molécule d'ADN sous forme de deux chaînes hélicoïdales enroulées autour d'un même axe et maintenues sous forme d'une structure à deux brins ou structure bicaténaire (du latin catena, « chaîne ») grâce à l'« appariement » des bases se faisant vis-à-vis sur l'une et l'autre chaîne (fig. 4) : les appariements sont toujours adénine-thymine (deux liaisons non covalentes ou liaisons hydrogène) et guanine-cytosine (trois liaisons non covalentes).
Cette structure qui réunit deux chaînes réciproquement complémentaires à l'égard de leurs séquences nucléotidiques est compatible avec la réplication de la molécule d'ADN (cf. infra).
On voit sur la figure 4b que les deux chaînes ont une polarité opposée : ce sont des chaînes antiparallèles ; de telle sorte que les extrémités 3′ et 5′ sont inversement placées dans chacun des deux brins. La complémentarité séquentielle et l'antiparallélisme des deux chaînes expriment le fait que leur structure est réciproquement codée (fig. 4c).
Pour mesurer la masse moléculaire des ADN, diverses méthodes ont été expérimentées avec plus ou moins de succès : diffusion, diffraction de la lumière, viscosité, sédimentation. En principe, la méthode qui offre la plus grande sécurité est celle qui consiste à mesurer effectivement la longueur de molécules entières sur un cliché pris au microscope électronique (fig. 5). Connaissant d'une part l'agrandissement et sachant d'autre part que l'unité de masse moléculaire par unité de longueur pour la chaîne de l'ADN est 1,92 × 106 par nanomètre, la masse moléculaire totale peut aisément être établie par un calcul simple, sachant que l'écart entre deux nucléotides consécutifs est de 0,34 nm (cf. fig. 4a). L'ultracentrifugation, effectuée sur des molécules natives, permet aussi d'obtenir d'assez bons résultats, et confirme ainsi les données précédentes.
On sait actuellement que la masse moléculaire de l'ADN monocaténaire du bactériophage ΦX-174 est de 1,6 × 106, ce qui est une très petite masse moléculaire pour un ADN contenant toute l'information nécessaire à la réplication d'un micro-organisme entier. En effet, l'ADN du virus de la vaccine comporte 240 000 paires de nucléotides, il a une masse moléculaire de 157 × 106. Quant à l'ADN d'un micro-organisme très connu comme E. coli, il comporte 3 400 000 paires de bases représentant une masse moléculaire de 2,3 × 109.
Accédez à l'intégralité de nos articles
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Jacques KRUH : docteur en médecine, docteur ès sciences, professeur de biochimie à la faculté de médecine de Cochin-Port-Royal
- Ethel MOUSTACCHI : directeur de recherche de classe exceptionnelle au C.N.R.S., unité 1292 (génotoxicologie et réparation de l'ADN)
- Michel PRIVAT DE GARILHE : ingénieur-docteur, docteur ès sciences, professeur au Conservatoire national des arts et métiers, ingénieur E.S.C.I.L.
- Alain SARASIN : directeur de recherche au C.N.R.S., directeur de l'Institut de recherche sur le cancer, agrégé de l'Université
Classification
Médias
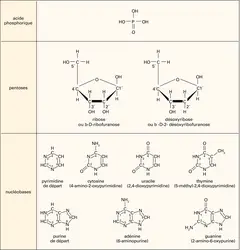
Molécules constitutives des nucléotides
Encyclopædia Universalis France
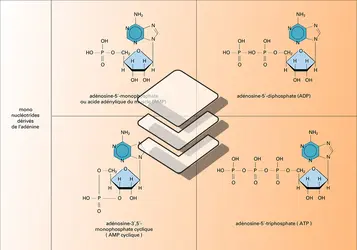
Nucléosides et nucléotides dérivés de l'adénine
Encyclopædia Universalis France
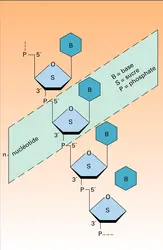
Polynucléotide
Encyclopædia Universalis France
Autres références
-
ADN (acide désoxyribonucléique) ou DNA (deoxyribonucleic acid)
- Écrit par Michel DUGUET , Encyclopædia Universalis , David MONCHAUD et Michel MORANGE
- 10 074 mots
- 10 médias
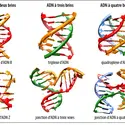 Le 25 avril 1953, l'Américain James D. Watson et le Britannique Francis H. C. Crick, récipiendaires, en 1962, avec le Britannique Maurice Wilkins du prix Nobel de physiologie ou médecine, proposaient, dans la célèbre revue scientifique anglaise Nature, une structure tridimensionnelle...
Le 25 avril 1953, l'Américain James D. Watson et le Britannique Francis H. C. Crick, récipiendaires, en 1962, avec le Britannique Maurice Wilkins du prix Nobel de physiologie ou médecine, proposaient, dans la célèbre revue scientifique anglaise Nature, une structure tridimensionnelle... -
ALTMAN SIDNEY (1939-2022)
- Écrit par Georges BRAM et Encyclopædia Universalis
- 377 mots
Le biochimiste américain d'origine canadienne Sidney Altman est né le 7 mai 1939 à Montréal (Canada). Après des études de physique au Massachusetts Institute of Technology (MIT) à Cambridge et une année à l'université Columbia à New York, il fait, à l'université de Boulder (Colorado), des recherches...
-
ANTIGÈNES
- Écrit par Joseph ALOUF
- 7 384 mots
- 5 médias
 L'immunogénicité des acides nucléiques, longtemps niée, est désormais bien établie, mais ne concerne que certains acides ribonucléiques (ARN) et désoxyribonucléiques (ADN). Les autoanticorps anti-ADN caractérisent certaines maladies auto-immunes (lupus érythémateux disséminé). Enfin, certains lipides...
L'immunogénicité des acides nucléiques, longtemps niée, est désormais bien établie, mais ne concerne que certains acides ribonucléiques (ARN) et désoxyribonucléiques (ADN). Les autoanticorps anti-ADN caractérisent certaines maladies auto-immunes (lupus érythémateux disséminé). Enfin, certains lipides... -
ARN (acide ribonucléique) ou RNA (ribonucleic acid)
- Écrit par Marie-Christine MAUREL
- 2 772 mots
- 2 médias
Faut-il voir dans une molécule biologique omniprésente dans toute structure cellulaire des êtres vivants, l'ARN (acide ribonucléique), la première étape de l'histoire de la vie ? Les ARN contemporains sont-ils les fossiles d'anciennes molécules ? Les voies métaboliques primordiales...
- Afficher les 39 références
Voir aussi
- INTERFÉRON
- ENZYMES
- ADN RÉPÉTITIF
- MATURATION DE L'ARN
- PROMOTEUR, biologie moléculaire
- GÈNES EN MOSAÏQUE
- HYDROGÈNE LIAISON
- MICROSATELLITES, génétique moléculaire
- ATAXIE-TÉLANGIECTASIE
- PHOTOSENSIBILITÉ, biologie
- MICROSCOPIE ÉLECTRONIQUE
- HYBRIDATION MOLÉCULAIRE
- PURINES
- ADN RÉPARATION DE L'
- LI-FRAUMENI SYNDROME DE
- HÉRÉDITAIRES MALADIES ou MALADIES GÉNÉTIQUES
- GUANINE
- NUCLÉOSIDES
- NUCLÉOTIDES
- ADN POLYMÉRASE
- CODON, biologie moléculaire
- ARN DE TRANSFERT ou ARNt
- ARN MESSAGER ou ARNm
- ARN POLYMÉRASE
- CYTOSINE
- ADÉNINE
- THYMINE
- RÉPLICATION, biologie moléculaire
- TRANSCRIPTION, biologie moléculaire
- TRADUCTION, biologie moléculaire
- POLYNUCLÉOTIDE
- URACILE
- COCKAYNE SYNDROME DE
- PYRIMIDINES
- STRUCTURE, biologie
- NUCLÉASES
- RAYONNEMENTS IONISANTS ou RADIATIONS IONISANTES
- CHLORURE DE CÉSIUM
- PHOSPHORIQUES ACIDES
- SÉQUENÇAGE, génétique moléculaire
- SÉPARATION, chimie
- GÈNE P53
- MASSE MOLÉCULAIRE
- PHOSPHORYLATION
- LIGASES
- ARN POLYMÉRASE ARN-DÉPENDANTE
- AVERY OSWALD THEODORE (1877-1955)
- GÉNOME
- BIOLOGIE MOLÉCULAIRE
- RIBOSE
- DÉSOXYRIBOSE
- HISTONES
- TRICHOTHIODYSTROPHIE
- DIVISION CELLULAIRE
- BIOSYNTHÈSES
- PROTÉINES BIOSYNTHÈSE DES
- INFORMATION GÉNÉTIQUE
- RECOMBINAISON GÉNÉTIQUE
- TRANSFORMATION BACTÉRIENNE
- TRANSDUCTION BACTÉRIENNE
- RÉGULATION GÉNÉTIQUE
- PATRIMOINE GÉNÉTIQUE
- EXPRESSION GÉNÉTIQUE
- EXON
- INTRON
- CONJUGAISON BACTÉRIENNE
- ARABINOSIDES
- ENZYMOPATHIES
- NUCLÉOTIDIQUE SÉQUENCE
- GÉNÉTIQUE MOLÉCULAIRE
- STAHL FRANKLIN WILLIAM (1929- )
- MOLÉCULES BIOLOGIQUES, structure et fonction
- MACROMOLÉCULES BIOLOGIQUES
- ANTICODON, génétique moléculaire
- ARN RIBOSOMIQUE ou ARNr
- CYTOTOXICITÉ
- CYCLE CELLULAIRE
- PEAU CANCER DE LA
- SPECTRE D'ABSORPTION
- PROTÉINES
- CÔLON CANCER DU
- ÉPISSAGE, génétique moléculaire
- MÉTHYLATION, biologie moléculaire
- MUTAGENÈSE
- TRANSCRIPTASE INVERSE ou REVERSE TRANSCRIPTASE
- DÉFICITS ENZYMATIQUE & MÉTABOLIQUE
- HÉLICASES
- BASES NUCLÉIQUES ou BASES AZOTÉES
- LYNCH SYNDROME DE
