- 1. Des organismes difficiles à définir
- 2. Le paradoxe d'organismes photosynthétiques non apparentés entre eux
- 3. Place des algues aujourd'hui dans le monde vivant
- 4. La théorie de l'endosymbiose et l'acquisition de la photosynthèse par les eucaryotes
- 5. L'évolution des eucaryotes et la dissémination de la photosynthèse : le résultat d'emboîtements successifs d'organismes
- 6. Conséquence de cette nature chimérique sur la définition des algues
- 7. Microalgues et macroalgues
- 8. Habitat
- 9. Utilisations, importance et nuisances
- 10. Bibliographie
ALGUES
Article modifié le
L'évolution des eucaryotes et la dissémination de la photosynthèse : le résultat d'emboîtements successifs d'organismes

Évolution des algues par emboîtements successifs d’organismes
Encyclopædia Universalis France

Certaines algues unicellulaires rouges ou vertes ont elles-mêmes été à l'origine de symbioses avec des groupes appartenant à des lignées autres que celle des Plantae (fig. 3). Possédant un plaste dérivé d'une cyanobactérie endosymbiotique, ces algues unicellulaires rouges et vertes sont devenues à leur tour des plastes et ont ainsi apporté la photosynthèse oxygénique chez ces lignées d'organismes hétérotrophes. Ces endosymbioses sont dites secondaires puisque les algues impliquées résultent déjà d'une endosymbiose (dite primaire) avec une cyanobactérie. Plusieurs événements de ce type se sont ainsi produits. Chez les euglénophycées, une endosymbiose secondaire a eu lieu entre une algue verte appartenant aux chlorophytes et l'ancêtre de ces euglénophycées. Un événement similaire s'est produit avec une chlorophyte, relativement proche mais distincte de celle à l'origine du plaste des euglénophycées, chez l'ancêtre des chlorarachniophycées. Chez les cryptophytes, les haptophytes, les algues brun-doré et, probablement, les dinophytes, une algue rouge symbiotique a évolué en plaste. Chez les cryptophytes et les chlorarachniophycées, les restes du cytoplasme et du noyau, respectivement d'une algue rouge (chez les cryptophytes) et d'une algue verte (chez les chlorarachniophycées), sont encore observables. Ce noyau relictuel, nommé nucléomorphe, contient encore un génome très réduit qui a pu être étudié : il recèle bien des chromosomes eucaryotiques, linéaires et pourvus de télomères, et non pas circulaires comme ceux des bactéries. Le compartiment contenant ces restes de cytoplasme est nommé compartiment périplastidial et, chez les cryptophytes, on y retrouve les grains d'amidons présents normalement dans le cytoplasme des algues rouges. Chez les haptophytes, les algues brun-doré et les dinophytes, le plaste est doré, bien que provenant d'une algue rouge, car les pigments rouges ont été perdus et remplacés par des xanthophylles (pigments proches du carotène). L'analyse des génomes de ces plastes d'origine endosymbiotique secondaire et des nucléomorphes a confirmé leur origine. Toutefois, la manière dont l'algue rouge a donné naissance aux plastes de ces organismes très divers reste encore mystérieuse. Les plastes des cryptophytes et des haptophytes, par exemple, présentent dans leur génome des caractéristiques uniques qui indiquent qu'ils ont une origine commune. Or ces deux groupes d'algues ne sont pas étroitement apparentés et ne présentent pas un ancêtre commun direct qui aurait pu s'associer avec l'algue rouge avant leur divergence. Il faut donc admettre que les cryptophytes et les haptophytes se sont associées séparément avec la même algue rouge, un événement hautement improbable, sauf si on admet que l'algue rouge s'est d'abord associée à l'ancêtre des cryptophytes (un flagellé hétérotrophe), puis qu'une cryptophyte pourvue d'un plaste dérivé d'une algue rouge s'est à son tour associée avec l'ancêtre des haptophytes (un autre flagellé hétérotrophe). Le plaste des haptophytes serait alors d'origine endosymbiotique tertiaire (un plaste dérivé d'une cryptophyte dont le plaste est lui-même dérivé d'une algue rouge dont le plaste est dérivé d'une cyanobactérie). Cette hypothèse est loin d'être improbable car les plastes des dinophytes sont d'origines multiples et des cas d'endosymbioses avec des haptophytes, des cryptophytes ou diverses ochrophytes sont bien démontrés chez les dinophytes. Chez les dinophytes, le plaste originel a souvent été perdu (la moitié environ des dinophytes en sont dépourvus) mais souvent aussi remplacé (par une algue verte, une algue brun-doré, une haptophyte ou une cryptophyte). Chez certaines dinophytes, le plaste a ainsi été perdu puis remplacé par une première diatomée, perdu à nouveau puis remplacé par une seconde diatomée ! Ces dinophytes, dont le plaste ancestral est au moins d'origine tertiaire et peut-être quaternaire, sont elles-mêmes en symbiose avec divers organismes marins, en particulier les coraux. Elles prennent alors le nom de zooxanthelles et les coraux ne peuvent survivre sans leurs algues symbiotiques. Des algues vertes ou des cyanobactéries s'associent avec des champignons pour former des lichens. Des cyanobactéries ou des algues vertes ou rouges s'associent aussi avec un grand nombre d'organismes aquatiques. La série des enchâssements ne s'arrête donc pas aux algues, lesquelles ont été qualifiées de « poupées russes » dans la littérature scientifique en raison des emboîtements successifs dont elles résultent. On a pu montrer aussi que certains sporozoaires ont un plaste qui n'est plus fonctionnel ; ces parasites sont donc des algues qui ont perdu leur plaste ou dont le plaste n'est plus capable d'effectuer la photosynthèse.
L' origine endosymbiotique des plastes et l'évolution des algues par emboîtements successifs expliquent comment des organismes sans lien de parenté directe entre eux ont acquis la possibilité de faire la même photosynthèse que les cyanobactéries. Puisque ces bactéries sont à l'origine des plastes des Plantae, on comprend bien que la photosynthèse de ces deux types très différents d'êtres vivants soit la même. Des microalgues vertes ou rouges (de nature végétale), contenant un plaste d'origine cyanobactérienne, ont ensuite établi des relations d'endosymbiose avec divers organismes (de nature non végétale) pour former des groupes d'algues non apparentés avec les autres et qui ont parfois à leur tour donné lieu à des relations endosymbiotiques pour former des lignées algales supplémentaires. La photosynthèse d'origine cyanobactérienne a ainsi été transportée d'association en association.
Accédez à l'intégralité de nos articles
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Bruno DE REVIERS : enseignant-chercheur au Muséum national d'histoire naturelle, Paris
Classification
Médias
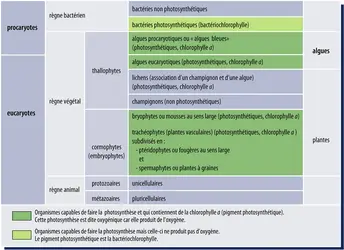
Ancienne classification du monde vivant en trois règnes
Encyclopædia Universalis France

Cyanobactérie : rivulaire (
B. De Reviers

Algue brune : fucus vésiculeux (
B. De Reviers
Autres références
-
AQUACULTURE
- Écrit par Lucien LAUBIER
- 11 442 mots
- 8 médias
 Plusde deux cents genres d'algues marines benthiques, correspondant à plusieurs milliers d'espèces, ont été étudiés en vue d'une exploitation. Une quinzaine d'espèces seulement figurent dans les statistiques de la F.A.O. Les différentes utilisations de ces végétaux peuvent être regroupées autour de...
Plusde deux cents genres d'algues marines benthiques, correspondant à plusieurs milliers d'espèces, ont été étudiés en vue d'une exploitation. Une quinzaine d'espèces seulement figurent dans les statistiques de la F.A.O. Les différentes utilisations de ces végétaux peuvent être regroupées autour de... -
BIOCARBURANTS ou AGROCARBURANTS
- Écrit par Jean-Paul CHARVET et Anthony SIMON
- 6 509 mots
- 10 médias
 La troisième génération est fondée sur l’utilisation d’algues (micro-algues ou macro-algues). Sa productivité à l’hectare serait de dix fois supérieure à celle de la première génération.
La troisième génération est fondée sur l’utilisation d’algues (micro-algues ou macro-algues). Sa productivité à l’hectare serait de dix fois supérieure à celle de la première génération. -
BIOTECHNOLOGIES
- Écrit par Pierre TAMBOURIN
- 5 368 mots
- 4 médias
 ...sont déjà utilisées dans différentes activités industrielles et pourraient être modifiées par voie moléculaire. Tel est le cas de polysaccharides issus d'algues (alginate, carraghénanes, etc.) qui servent de stabilisant, gélifiant ou épaississant. Tous ces produits sont omniprésents dans la vie quotidienne...
...sont déjà utilisées dans différentes activités industrielles et pourraient être modifiées par voie moléculaire. Tel est le cas de polysaccharides issus d'algues (alginate, carraghénanes, etc.) qui servent de stabilisant, gélifiant ou épaississant. Tous ces produits sont omniprésents dans la vie quotidienne... -
CARBURANTS POUR L'AVIATION
- Écrit par Paul NASH et Odile PÉTILLON
- 3 244 mots
- 3 médias
 – Les micro-algues. Celles-ci se développent plus rapidement que des plantes terrestres, contiennent des quantités intéressantes d'huile et peuvent être cultivées dans des eaux salées ou saumâtres (biocarburants de troisième génération).
– Les micro-algues. Celles-ci se développent plus rapidement que des plantes terrestres, contiennent des quantités intéressantes d'huile et peuvent être cultivées dans des eaux salées ou saumâtres (biocarburants de troisième génération). - Afficher les 53 références
Voir aussi
- EUCARYOTES
- CHLOROPHYTES
- CHLOROPHYCOPHYTES ou ALGUES VERTES
- RHODOPHYTES ou ALGUES ROUGES
- XANTHOPHYLLE
- DINOPHYTES ou DINOFLAGELLÉS ou PÉRIDINIENS
- ZOOXANTHELLES
- PROCARYOTES ou PROTOCARYOTES
- AQUATIQUE VIE
- BENTHIQUE VIE
- PHYLOGÉNIE MOLÉCULAIRE
- ENDOSYMBIOTIQUE THÉORIE
- GLAUCOPHYTES
- CRYPTOPHYTES, phycologie
- CHLORARACHNIOPHYCÉES
- HAPTOPHYTES
- OCHROPHYTES ou ALGUES BRUN-DORÉ
