
ARCHÉOBACTÉRIES ou ARCHÉES
Article modifié le
Diversité des archées
Les méthanogènes
Les archées méthanogènes sont les premières à avoir été identifiées en tant que telles par Carl Woese. Ce sont des anaérobies strictes qui utilisent le dioxyde de carbone ou d'autres composés organiques simples, et l'hydrogène, pour produire du méthane, un processus appelé méthanogenèse. Ce métabolisme, qui semble fait sur mesure pour l'atmosphère terrestre primitive, explique le choix initial fait par Carl Woese du terme archéobactérie. La méthanogenèse est en fait un mécanisme très complexe qui met en jeu des coenzymes spécifiques et qui n'existe que chez les archées. Les archées méthanogènes sont présentes dans tous les biotopes anaérobies (marécages, rizières, intestins des animaux, sources hydrothermales terrestres et sous-marines) et peuvent vivre dans toute la gamme de température compatible avec la vie, de 0 0C (psychrophiles) à 110 0C (hyperthermophiles de l'ordre des Méthanopyrales). Les archées méthanogènes sont présentes en abondance dans notre intestin (en particulier l'espèce Methanobrevibacter smithii), et elles ont été récemment associées à des pathologies gingivales au sein de « consortiums bactériens ». Les méthanogènes ont des morphologies très variées (coques, bacilles, sarcines) et peuvent être à Gram positif ou négatif. Les méthanogènes à Gram positif (ordre des Méthanobactériales) possèdent dans leur paroi de la pseudomuréine qui ressemble à la muréine des parois bactériennes (dans les deux cas, il s'agit de longues chaînes polysaccaridiques reliées entre elles par de courtes chaînes peptidiques), tout en présentant des différences majeures (par exemple les acides aminés des ponts peptidiques sont de forme L chez les bactéries et D chez les Méthanobactériales).
Les halophiles
Les archées halophiles (qui aiment le sel) sont des micro-organismes aérobies stricts dont la particularité est de vivre dans les milieux proches de la saturation en sel (lacs salés, marais salants). Ils maintiennent leur pression osmotique interne en accumulant des concentrations équivalentes de chlorure de potassium dans leur cytoplasme, ce qui implique l'existence d'enzymes capables de fonctionner à des concentrations de sel tout à fait inhabituelle. Les archées halophiles, qui vivent en surface, se protègent des rayons ultraviolets par la présence de pigments colorés (bactériorubérine), d'où leur couleur rouge. Certaines halophiles possèdent de plus une « membrane pourpre » composée de bactériorhodopsine, une protéine responsable d'un mécanisme de photosynthèse non chlorophyllienne. Les archées halophiles ont des morphologies très variées – coque pour les Halococcus, bacille pour les Halobacterium, forme plus ou moins géométrique pour les Haloarcula. Les cellules des Haloquadrata, récemment cultivées, sont les seules cellules parfaitement carrées (en coupe) connues dans le monde vivant.
Les thermoacidophiles
Les thermoacidophiles se divisent en deux grands groupes, les Thermoplasmatales, qui sont des thermophiles modérées (température optimale de croissance entre 40 et 60 0C) mais qui peuvent être des acidophiles extrêmes (Picrophilus torridus pousse à pH 0 !) et les Sulfolobales, qui sont des thermophiles extrêmes (température optimale de croissance entre 70 et 80 0C) et qui pousse dans une gamme de pH allant de 3 à 4. Les thermoacidophiles sont généralement aérobies ou micro-aérobies, mais certains Sulfolobus peuvent être anaérobies. Ces archées vivent dans les sources chaudes terrestres acides. Sulfolobus solfataricus est l'une des archées modèles les plus étudiées par les biochimistes et les biologistes moléculaires.
Les hyperthermophiles
Les hyperthermophiles sont, par définition, les archées dont la température optimale de croissance est supérieure ou égale à 80 0C (définition donnée par le microbiologiste allemand Karl Stetter qui est à l'origine de la description de la plupart de ces micro-organismes). Ces archées vivent dans les sources chaudes terrestres ou marines et dans les parois des cheminées hydrothermales sous-marines. L'hyperthermophile qui présente la température maximale de croissance la plus élevée, 113 0C, est Pyrolobus fumarii. Pour le moment, tous les micro-organismes connus capables de vivre à des températures supérieures à 95 0C font partie du domaine des archées, car les records de thermophilie pour les bactéries et les eucaryotes sont respectivement de 95 0C (Aquifex pyrophilus) et 62 0C (des champignons détectés à Yellowstone). La plupart des archées hyperthermophiles sont des anaérobies strictes, mais quelques-unes sont microaérobies (Pyrobaculum aerophilum, Aeropyrum pernix). De nombreux hyperthermophiles sont chimio-lithoautotrophes (genres Archaeoglobus...) et utilisent l'énergie de l'hydrogène pour réduire différents métaux ou composés inorganiques (CO2, S, SO42— NO3-, Fe). D'autres, tels les Thermococcales, sont hétérotrophes. La plupart nécessitent la présence de soufre dans leur milieu de culture et produisent de l'hydrogène sulfureux (H2S). L'une des plus petites cellules connues dans le monde vivant est une archée hyperthermophile de très petite taille, Nanoarchaeum equitans, qui vit en symbiose avec une autre archée hyperthermophile, Ignicoccus islandicus. Les lipides géants des archées, capables de former des monocouches lipidiques (caldarchaeol), ont sans doute joué un rôle important dans la capacité de celles-ci à coloniser les biotopes les plus chauds, car les membranes formées de lipides classiques deviennent perméables à haute température, ce qui empêche la formation des gradients de protons ou d'autres ions nécessaires à la synthèse d'ATP. Un autre facteur qui a sans doute permis à certaines archées d'évoluer vers le caractère hyperthermophile est l'acquisition d'une enzyme particulière, la reverse gyrase, qui augmente fortement le nombre de liens topologiques entre les deux brins de la molécule d'ADN. Cette protéine est la seule qui soit spécifique des hyperthermophiles, son rôle précis in vivo restant encore inconnu. Contrairement aux protéines classiques, les protéines des hyperthermophiles sont très stables à haute température ; certaines peuvent être autoclavées plusieurs heures à 120 0C sans perdre leur activité. La comparaison entre des protéines mésophiles et hyperthermophiles a montré que cette stabilité était due à une augmentation du nombre d'interactions faibles, en particulier des interactions ioniques entre les acides aminés à la surface de la protéine. Les protéines des hyperthermophiles sont souvent plus faciles à cristalliser que leurs homologues mésophiles (des milieux tempérés) et ont été prises comme objets d'étude privilégiés dans de nombreux programmes de génomique structurale. Les ARN de transferts et les ARN ribosomaux des hyperthermophiles sont stabilisés par une proportion élevée de paires de bases GC (lesquelles sont reliées par trois liaisons hydrogènes, contre deux pour les paires de bases AT) et la présence de nombreux nucléotides modifiés qui permettent la formation d'interactions stabilisatrices supplémentaires. Le génome des hyperthermophiles n'est pas spécialement riche en paires de bases GC car l'ADN intracellulaire est protégé contre la dénaturation par les liens topologiques entre les deux brins de l'hélice. Par contre, l'ADN des hyperthermophiles doit être protégé à haute température contre la dégradation chimique par des mécanismes de réparation particulièrement efficaces mais encore peu explorés.
Les archées qui oxydent l'ammoniaque
Dans les années 1990, de nouveaux groupes d'archées mésophiles ou psychrophiles ont été identifiés dans des biotopes tempérés ou froids, grâce aux méthodes de l'écologie moléculaire microbienne, qui permettent de détecter un organisme in situ, sans avoir à le cultiver, en amplifiant son ARNr 16S par la méthode de polymérisation en chaîne (PCR). Ces archées peuvent être très abondantes, formant par exemple jusqu'à 25 p. 100 de l'ensemble de la biomasse océanique. Des travaux de métagénomique (isolement direct de gènes dans l'environnement) et la culture de l'un de ces organisme en 2005 ont mis en évidence parmi ces archées mésophiles un grand groupe de micro-organismes capables d'oxyder l'ammoniaque. Des travaux récents indiquent que ces archées pourraient être les principaux acteurs du cycle de l'azote dans les océans et dans le sol, rôle que l'on réservait jusque-là à un groupe particulier de bactéries.
Accédez à l'intégralité de nos articles
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Patrick FORTERRE : professeur émérite à l'université Paris-Saclay, professeur honoraire à l'Institut Pasteur
Classification
Autres références
-
ARCHÉES ASGARD
- Écrit par Patrick FORTERRE
- 3 786 mots
- 3 médias
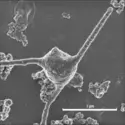
Les Archées, l’un des trois domaines du monde vivant – les deux autres étant les Bactéries et les Eucaryotes –, sont des micro-organismes unicellulaires et dépourvus de noyau extrêmement diversifiés. Leur mise en culture permet de les étudier et de les caractériser, montrant qu’elles présentent...
-
EXOBIOLOGIE
- Écrit par Vassilissa VINOGRADOFF
- 8 001 mots
- 4 médias
 Lesarchées (anciennement appelées archéobactéries) intéressent particulièrement les biologistes. Ces microorganismes procaryotes, particulièrement adaptés aux environnements extrêmes (en termes de pression, température, salinité, nutriments…), sont morphologiquement semblables aux bactéries...
Lesarchées (anciennement appelées archéobactéries) intéressent particulièrement les biologistes. Ces microorganismes procaryotes, particulièrement adaptés aux environnements extrêmes (en termes de pression, température, salinité, nutriments…), sont morphologiquement semblables aux bactéries... -
MICROBIOLOGIE
- Écrit par Corinne DOREL , Philippe LEJEUNE et Jean-Michel PANOFF
- 3 881 mots
- 9 médias
 Les archéobactéries vivent dans des milieux extrêmement défavorables et ont des caractéristiques si spécifiques tant au niveau de leur expression génétique que de leur métabolisme que certains biologistes considèrent que le monde vivant est composé de trois domaines : les eucaryotes, les archéobactéries...
Les archéobactéries vivent dans des milieux extrêmement défavorables et ont des caractéristiques si spécifiques tant au niveau de leur expression génétique que de leur métabolisme que certains biologistes considèrent que le monde vivant est composé de trois domaines : les eucaryotes, les archéobactéries... -
OCÉAN ET MERS (Vie marine) - Vie dans les grandes profondeurs
- Écrit par Lucien LAUBIER
- 3 870 mots
- 2 médias
...présentes dans les fluides hydrothermaux, sur les substrats basaltiques et les tubes des vestimentifères et des polychètes ; il s'agit principalement d' archæbactéries qui constituent, à côté des bactéries vraies ou procaryotes et des eucaryotes, un troisième règne caractérisé notamment par les molécules... -
ORGANISME VIVANT
- Écrit par Jean GÉNERMONT
- 1 651 mots
...rapidement diversifiée, avec pour résultat l'individualisation de lignées dont trois seulement se sont maintenues jusqu'à nos jours. Dans deux d'entre elles, archées et eubactéries, qui diffèrent par une foule de caractères moléculaires, la cellule, dite procaryote, n'est pas compartimentée, d'où notamment... - Afficher les 7 références
Voir aussi
