AUXINES
Article modifié le
Homéostasie de l'auxine
Les réponses contrôlées par l'auxine dépendent de sa concentration locale dans un tissu donné, à un instant donné. Les ajustements constants de concentrations en auxine résultent de l'équilibre entre plusieurs processus, à savoir biosynthèse, conjugaison, compartimentation, dégradation et transport à courtes ou longues distances.
Biosynthèse de l'AIA
Chez les plantes supérieures, les étapes conduisant à la synthèse de l'AIA ne sont pas encore toutes élucidées. Ce n'est que depuis les années 1980 que des éléments précis concernant les voies de biosynthèse de l'AIA commencent à émerger. Les données viennent pour une part d'analyses biochimiques utilisant des marquages isotopiques qui ont permis l'identification de composés intermédiaires de synthèse, et d'autre part d'approches génétiques amenant au clonage de gènes codant des enzymes de biosynthèse de l'AIA.
Les données biochimiques supportent l'hypothèse de deux voies distinctes de biosynthèse, l'une dépendant du tryptophane et l'autre non. Elles dérivent toutefois toutes les deux de l'anthranilate, précurseur du tryptophane, mais la voie indépendante du tryptophane dériverait directement de l'indole-3-glycérol-phosphate ou de l'indole, tous deux précurseurs du tryptophane. En ce début de xxie siècle, seuls des gènes codant certaines enzymes de biosynthèse de la voie dépendant du tryptophane ont été identifiés.

La synthèse d'AIA se produit plutôt dans les tissus jeunes, en particulier dans les très jeunes feuilles, mais aussi dans les cotylédons et les racines. Sur la plante modèle Arabidopsis thaliana (L.) Heynh. (arabette des dames, famille des Brassicaceae ou Crucifères), la distribution et la biosynthèse de l'AIA ont été étudiées au cours du développement végétatif de la plante. Les très jeunes feuilles, de moins de 0,5 mm de long, renferment environ 250 pg d'AIA par milligramme de poids frais et leur capacité de synthèse est très élevée. Le contenu en AIA de feuilles ayant terminé leur croissance est plus de cent fois inférieur et leur capacité de synthèse a considérablement décru.
L'auxine est également synthétisée par des micro-organismes, parmi lesquels de nombreuses bactéries du sol, phytopathogènes (Agrobacterium ou Pseudomonas) ou non (Rhizobium). Différentes voies, dépendantes ou non du tryptophane, ont été rapportées selon les micro-organismes. La production d'auxine par les bactéries phytopathogènes interfère avec le métabolisme des hormones de la plante infectée. La formation de tumeurs, correspondant à une prolifération cellulaire accrue consécutive à l'infection par certains pathogènes, est une réponse à l'accroissement de concentration en auxine produite par les bactéries.
Métabolisme
Le métabolisme de l'AIA recouvre des processus d'oxydation et de conjugaison qui modifient soit le noyau indole soit la chaîne latérale et causent la perte de l'activité biologique. Certaines formes de conjugaison font partie du catabolisme de l'AIA, dans la mesure où les conjugués formés ne peuvent être retransformés en AIA actif.
Le catabolisme décarboxylatif de l'AIA implique aussi bien des modifications du noyau indole que de la chaîne latérale. Ces réactions complexes sont catalysées par une variété d'enzymes de la classe des peroxydases, généralement appelées des AIA-oxydases. Les produits de dégradation sont peu abondants dans les extraits de plantes et il reste difficile d'évaluer l'importance relative de la dégradation par décarboxylation. L'autre voie de catabolisme de l'AIA est l'oxydation sans décarboxylation du noyau aromatique. L'oxydation intervient soit directement sur l'AIA libre soit sur un conjugué aspartate (AIAsp). Ces deux mécanismes coexistent et leur importance varie selon les plantes.
Pendant la conjugaison, la chaîne latérale est modifiée ; deux types de conjugués peuvent être formés : des conjugués de type ester, où le groupement carboxyle de la chaîne latérale est lié par un pont oxygène à des sucres (par exemple du glucose) ou des polyols cycliques (comme l'inositol) ; ou des conjugués de type amide où le groupement carboxyle forme une liaison amide avec des acides aminés (-Ala, -Glu, -Leu, -Asp) ou des polypeptides.
La conjugaison de l'AIA est un moyen efficace de contrôler la concentration intracellulaire en AIA libre dans les cellules. Lors d'un traitement de cellules en culture par une source exogène d'AIA, une part importante de l'AIA intracellulaire est en quelque sorte « neutralisée » par l'accumulation de conjugués. Ce mécanisme est très rapide. Certaines auxines synthétiques peuvent également être rapidement conjuguées, comme le 1-ANA. En revanche, d'autres auxines synthétiques, comme par exemple le 2,4-D, ne sont que très difficilement métabolisées.
Transport de l'auxine
L'AIA est un acide faible dont le pKa est d'environ 4,8. L'équilibre entre la forme protonée AIAH et la forme anionique AIA— évolue donc en fonction du pH environnant. Ainsi, à pH neutre ou basique, la forme anionique l'emporte alors qu'à pH acide, l'équilibre se déplace en faveur de la forme protonée. Celle-ci est lipophile et facilement diffusible au travers de la membrane plasmique alors que l'anion ne diffuse pas. Le pH moyen de l'espace apoplastique (espace extracellulaire compris entre la membrane plasmique et la paroi) est d'environ 5,5. Dans ces conditions, une fraction significative de l'AIA est sous forme protonée, ce qui favorise son entrée dans la cellule, à la faveur du gradient de pH. Dans le cytosol dont le pH est d'environ 7,4, l'AIAH est aussitôt dissocié en AIA— et H+.
L'AIA est aussi transporté au travers de la membrane plasmique par des transporteurs sélectifs, qui contrôlent soit son entrée (transporteurs d'influx), soit sa sortie (transporteurs d'efflux). Les protéines impliquées dans le transport d'influx, identifiées par des études génétiques, appartiennent à une famille de perméases aux acides aminés, nommées AUX1/LAX, du nom des mutants d'Arabidopsis thaliana isolés initialement aux1 (auxin resistant 1) et lax (like aux1). Les transporteurs d'influx ne sont pas présents dans toutes les cellules et semblent être associés au chargement de l'AIA vers le phloème, et à des redistributions localisées d'AIA nécessaires à la croissance et au développement des plantes (par exemple la formation des racines latérales ou le gravitropisme). Le transport d'efflux de l'AIA apparaît plus complexe et fait intervenir plusieurs types de protéines. Ces transporteurs d'efflux jouent un rôle essentiel dans le contrôle de l'homéostasie, l'établissement de gradients de concentration localisés et le transport à longue distance de l'AIA, des lieux de synthèse situés dans la région apicale vers les racines. La capacité de redistribution de l'AIA, étroitement liée à l'activité et à la localisation des transporteurs, agit tel un moteur du développement de la plante. C'est un système dynamique, en constante évolution et adaptation. La vitesse moyenne de transport polarisé de l'AIA dans les tiges oscille entre 5 et 20 mm par heure selon les plantes. À ce transport actif s'ajoute un transport par le phloème.
Accédez à l'intégralité de nos articles
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Catherine PERROT-RECHENMANN : directrice de recherche au C.N.R.S.
Classification
Médias

Phototropisme
Cathlyn Melloan/ Stone/ Getty Images
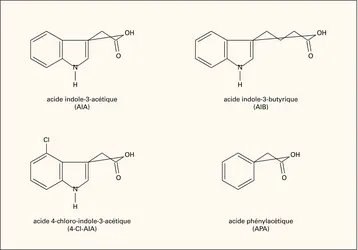
Auxines : structure chimique
Encyclopædia Universalis France
Autres références
-
PREMIÈRE HORMONE VÉGÉTALE : L'AUXINE
- Écrit par Claude LANCE
- 212 mots
En isolant, en 1928, à partir du coléoptile d'avoine (gaine entourant les jeunes feuilles au début de la germination), une substance (ultérieurement nommée auxine et identifiée à l'acide β-indolylacétique ou AIA) qui est capable de promouvoir l'élongation de cet organe, Fritz W. Went (1903-1990)...
-
COMMUNICATION CELLULAIRE
- Écrit par Yves COMBARNOUS
- 6 593 mots
- 7 médias
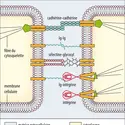 Ainsi,la balance des actions stimulantes de l’auxine (acide indole-acétique, IAA) et inhibitrices des cytokinines détermine le phénotype racinaire des cellules et la différenciation des racines. Les brassinostéroïdes et les gibbérellines superposent leurs actions à l’équilibre auxine-cytokinines...
Ainsi,la balance des actions stimulantes de l’auxine (acide indole-acétique, IAA) et inhibitrices des cytokinines détermine le phénotype racinaire des cellules et la différenciation des racines. Les brassinostéroïdes et les gibbérellines superposent leurs actions à l’équilibre auxine-cytokinines... -
CROISSANCE, biologie
- Écrit par Encyclopædia Universalis , André MAYRAT , Raphaël RAPPAPORT et Paul ROLLIN
- 14 766 mots
- 7 médias
 L' auxine, ou acide β-indolylacétique (AIA), qui est synthétisée dans les bourgeons et qui stimule la croissance des tiges, présente un transport polarisé ; on peut donc la considérer comme une hormone. Elle agit sur l'élongation des cellules dérivées de l'activité des méristèmes primaires. Son...
L' auxine, ou acide β-indolylacétique (AIA), qui est synthétisée dans les bourgeons et qui stimule la croissance des tiges, présente un transport polarisé ; on peut donc la considérer comme une hormone. Elle agit sur l'élongation des cellules dérivées de l'activité des méristèmes primaires. Son... -
DÉVELOPPEMENT, biologie
- Écrit par Georges DUCREUX , Hervé LE GUYADER et Jean-Claude ROLAND
- 19 226 mots
- 14 médias
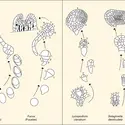 ...des explants qui n'en comportent pas, comme des portions d'entre-nœuds ou de feuilles. La réussite de cette opération exige la séparation de la plante mère et la réactivation ou la dédifférenciation des portions mises en culture sous l'influence d'activateurs de croissance, auxines et/ou cytokinies.
...des explants qui n'en comportent pas, comme des portions d'entre-nœuds ou de feuilles. La réussite de cette opération exige la séparation de la plante mère et la réactivation ou la dédifférenciation des portions mises en culture sous l'influence d'activateurs de croissance, auxines et/ou cytokinies. -
FRUITS
- Écrit par Encyclopædia Universalis , Jean-Luc REGNARD et Roger ULRICH
- 7 963 mots
- 13 médias
 Le développement des fruits charnus fait intervenir des substances de croissance ( auxines, gibbérellines, cytokinines, le plus souvent associées), comme l'ont montré les cultures de tissus de fruits.
Le développement des fruits charnus fait intervenir des substances de croissance ( auxines, gibbérellines, cytokinines, le plus souvent associées), comme l'ont montré les cultures de tissus de fruits. - Afficher les 8 références
Voir aussi
- TRANSPORTS MEMBRANAIRES
- VERGER
- HERBICIDES ou DÉSHERBANTS
- VÉGÉTALE BIOLOGIE
- ORGANOGENÈSE
- GRADIENT, biologie
- BOUTURAGE
- DÉVELOPPEMENT VÉGÉTAL ou ONTOGENÈSE VÉGÉTALE
- WENT FRITZ WARMOLT (1903-1990)
- DÉFOLIANTS
- RÉGULATION BIOLOGIQUE
- PHOTOTROPISME
- PERMÉASES
- TUMEURS VÉGÉTALES
- DIVISION CELLULAIRE
- BIOSYNTHÈSES
- DÉDIFFÉRENCIATION CELLULAIRE
- CYTOKININES
- AVENA TEST
- PARTHÉNOCARPIE
- INDOLE-3-ACÉTIQUE ACIDE (AIA)
- PHLOÈME ou LIBER, botanique
- TRYPTOPHANE
- PHYSIOLOGIE VÉGÉTALE ou PHYTOBIOLOGIE
- RÉGULATIONS BIOCHIMIQUES
- COLÉOPTILE, botanique
- BIOLOGIE HISTOIRE DE LA
