BASIDIOMYCÈTES
Article modifié le
Ce groupe de champignons rassemble la majeure partie des espèces aux fructifications de grande taille, celles qu'on va ramasser dans les bois ou dans les prés ; il comprend aussi des organismes microscopiques parasites de plantes ; ces derniers provoquent alors des maladies dont les noms (« rouilles », « charbons », etc.) évoquent la couleur des pustules que forment leurs fructifications sur les organes végétaux qu'ils ont envahis. Tous possèdent néanmoins une caractéristique commune : celle de produire leurs spores postméiotiques, ou basidiospores, à l'apex de diverticules (stérigmates) émis par la cellule mère, appelée baside.
Ce sont, en général, des organismes relativement peu compétitifs : aussi se réfugient-ils souvent dans ces sortes de « niches » que constituent le parasitisme, la symbiose ou la lignivorie, autant de modes de vie qui leur procurent des substrats dans lesquels ils ne se heurtent pas à la concurrence des Ascomycètes (ou de leurs formes imparfaites) et des Bactéries. La vie parasitaire est pratiquement de règle chez ceux des Basidiomycètes qui n'édifient pas de basidiocarpes agrégés volumineux (Urédinales, Ustilaginales, Exobasidiales). Les représentants des autres groupes sont, au contraire, plutôt saprophytiques ou symbiotiques.

Les espèces lignivores ont un comportement saprophytique, même lorsqu'elles se développent chez des arbres encore vivants : elles ne dégradent en réalité que des tissus morts (bois de cœur) et, en règle générale, seulement les parois cellulaires. Les autres saprophytes vivent le plus souvent aux dépens de débris végétaux peu décomposés, riches en éléments ligno-cellulosiques : les Basidiomycètes sont les seuls organismes capables de réaliser les premières étapes de leur dégradation.
Les Basidiomycètes participent également à la symbiose lichénique, certainement avec une fréquence bien supérieure à celle qu'on leur a longtemps attribuée. Toutefois, la mycorhization demeure pour eux le principal mode d'association symbiotique.
Dans le cycle des Basidiomycètes, le stade haploïde représente souvent une phase mineure, parfois même négligeable. Il ne se termine pas par une fécondation classique, telle qu'on l'observe chez les animaux ou les autres végétaux, mais par une simple fusion cytoplasmique (« plasmogamie » semblable à celle des Ascomycètes), sans fusion des noyaux. Ces derniers restent autonomes mais se divisent par des mitoses synchrones : chaque article est donc binucléé. Ce stade binucléé, dit « dicaryotique », s'achève à son tour au niveau de la baside, siège de la fusion des noyaux (« caryogamie ») et de la méiose.
On répartit habituellement les Basidiomycètes en trois grands ensembles : les Téliobasidiomycètes, chez qui la baside, souvent cloisonnée, naît de la germination d'une cellule à paroi épaisse (probaside) fonctionnant en propagule ou intégrée à une propagule (téleutospores pluricellulaires, regroupant plusieurs probasides), les Phragmobasidiomycètes, groupe assez hétérogène qui occupe une position quelque peu intermédiaire : la baside, toujours cloisonnée, peut être issue d'une probaside (mais alors non libérée, germant sur place) ou d'un apex hyphal, et les Holobasidiomycètes, dont la baside, non divisée, résulte de la différenciation directe d'un apex d'hyphe à paroi mince.
Les Téliobasidiomycètes
Les Téliobasidiomycètes sont des champignons parasites de plantes diverses. Parmi eux, les Urédinales, à cycle complexe, sont responsables des maladies dénommées « rouilles » car leurs fructifications constituent souvent, sur les organes envahis, des taches de couleur jaune, orangé, brun rouillé ou brun chocolat ; les Ustilaginales, au cycle beaucoup plus simple, provoquent les maladies appelées « charbons » ou « caries », selon que les amas noirs de leurs spores sont superficiels à maturité ou demeurent inclus dans les tissus de l'organe envahi.
Les Urédinales
Les Urédinales attaquent surtout les feuilles ou les rameaux de leurs hôtes, plus rarement les fleurs ou les fruits ; chez les arbres, leur développement peut entraîner des déformations de branches (balais de sorcières, etc.). Leur mycélium croît entre les cellules, dans la lamelle mitoyenne, envoyant des suçoirs de part et d'autre, dans le cytoplasme des cellules vivantes. Leurs spores sont formées à l'extrémité d' hyphes longues qui sortent par les stomates ou, plus souvent, sur des hyphes courtes rassemblées dans des réceptacles ou des pustules, d'abord sous-épidermiques, puis saillants à maturité par rupture de l'épiderme ; seules, les téleutospores peuvent, dans certains cas, rester internes pour n'être libérées qu'après la décomposition des tissus de l'hôte.
Leur cycle peut se dérouler entièrement sur des plantes de la même espèce (Urédinales autoïques) ou impliquer (Urédinales hétéroïques) des hôtes appartenant à deux espèces distinctes, voire très différentes : fougère et résineux, ou encore résineux et plante dicotylédone herbacée, par exemple. Dans ce dernier cas, une des espèces hôtes héberge la phase haploïde et supporte, après la plasmogamie, le premier type de spores dicaryotiques, les écidiospores. La seconde espèce hôte entretient l'ensemble de la phase dicaryotique, qui produit une ou plusieurs générations de spores de multiplication végétative (urédospores) puis les téleutospores, propagules mères des basides. Lorsqu'un, au moins, de ces types de spores manque, l'Urédinale est dite « microcyclique ».
La germination d'une basidiospore d'Urédinale donne naissance à un thalle haploïde qui peut édifier, à la surface de l'organe parasité, des petits conceptacles appelés pycnies ou spermogonies au sein desquels sont émises les spermaties haploïdes qui assurent, au moins dans certains cas, le rôle de gamètes mâles : libérées, elles peuvent réaliser la plasmogamie avec des « hyphes flexueuses » de la même spermogonie ou d'une autre, alors située ou non sur la même plante hôte. Indépendamment de la présence ou de l'absence de spermogonies, la plasmogamie peut aussi résulter de fusions entre hyphes végétatives à l'intérieur de l'organe hôte. Dans tous les cas, il en naît un dicaryon qui produit alors la première génération de spores dicaryotiques, les écidiospores.
Formées dans les « écidies » d'abord sous-épidermiques, les écidiospores, unicellulaires, sont émises successivement, en files serrées, par leurs cellules mères qui tapissent l'intérieur du réceptacle. Dans les cas les plus banals (type Aecidium), l'écidie est en forme de bourse globuleuse, délimitée par une collerette membraneuse (péridium) qui se déchire à maturité. Le péridium est parfois très allongé, s'ouvrant alors par des déchirures plus ou moins longitudinales et latérales ou localisées à proximité du sommet (type Roestelia, caractéristique du genre Gymnosporangium). Chez le type Caeoma, le péridium manque complètement, laissant l'écidie nue ou, chez le genre Phragmidium, seulement entourée de quelques filaments. Enfin, le type Peridermium réunit les écidies quelque peu naviculaires à péridium plus ou moins vésiculeux et dont les modes de déhiscence sont variables.
Quel que soit le type de la fructification écidienne, ce sont les écidiospores qui assurent, chez les Urédinales hétéroïques, le passage du parasite de son hôte écidien à l'hôte urédien et télien. Le mycélium dicaryotique issu de leur germination envahit les tissus du nouvel hôte et différencie des sores sous-épidermiques qui vont émerger : les urédosores, rassemblant des spores dicaryotiques unicellulaires, les urédospores, portées chacune par un court pédicelle ; l'ensemble n'est pas protégé par un péridium mais seulement, dans quelques cas, par une couronne de filaments périphériques. Ce stade urédien, qui peut se répéter plusieurs fois, représente une période d'intense multiplication végétative, permettant de nombreuses contaminations secondaires et le développement de l'épidémie au sein des populations de plantes hôtes.
En fin de saison, dans des sores indépendants des urédosores ou dans ces derniers et alors en mélange avec des urédospores, ou encore, parfois, à l'intérieur des tissus envahis, sont produites des spores d'un nouveau type, les téleutospores, organes de résistance à paroi épaisse, unicellulaires et nommées probasides ou pluricellulaires, chaque loge étant alors une probaside. À sa germination, qui a souvent lieu au printemps, la probaside engendre un court mycélium tétracellulaire (promycélium), véritable baside cloisonnée puisque chacun de ses quatre articles émet une basidiospore haploïde à l'extrémité d'un stérigmate. Plus rarement (genre Coleosporium), la téleutospore ne germe pas en un promycélium mais se cloisonne intérieurement pour délimiter quatre loges émettant chacune un stérigmate porteur d'une basidiospore. Ce sont les basidiospores, haploïdes, qui assurent le retour du parasite sur son hôte écidien.
Outre sa complexité, le cycle des Urédinales montre une originalité remarquable : celle d'une phase dicaryotique à la fois inféodée à la phase haploïde (développement de l'écidie, greffée sur le thalle haploïde comme l'est la fructification des Ascomycètes) et s'affranchissant de cette phase, souvent avec changement d'hôte, pour vivre de manière autonome. Chez tous les autres groupes de Basidiomycètes, au contraire, la phase dicaryotique mène une vie entièrement indépendante de la phase haploïde.
Les Ustilaginales
Comme les Urédinales, les Ustilaginales sont des parasites internes de plantes ; mais leur cycle est beaucoup plus simple et possède, en général, un stade saprophytique. Typiquement, la germination de leurs basidiospores donne naissance à des organismes haploïdes, saprophytes, quelque peu filamenteux ou, plus souvent, levuriformes. La plasmogamie s'effectue entre filaments ou cellules levuriformes, ou même parfois entre basidiospores voisines avant qu'elles ne se détachent de leurs stérigmates. Le mycélium dicaryotique est toujours parasite, infectant l'hôte au moment de la germination, au niveau des apex actifs ou à celui des inflorescences, restant selon les cas localisé au lieu de l'attaque ou inféodé aux zones méristématiques qu'il suit tout au long de leur croissance ; dans ce dernier cas, il peut alors se maintenir, plusieurs années, dans des organes à croissance pérenne (rhizomes de monocotylédones, par exemple).
La dicaryophase des Ustilaginales se termine par une désarticulation des hyphes mycéliennes, chaque article se différenciant en propagule à paroi épaisse, chargée de pigment noir. Ce sont autant de probasides dont la germination, accompagnée de la caryogamie et de la méiose comme chez les Urédinales, se traduit par l'émission d'un promycélium producteur des basidiospores. Le promycélium peut être cloisonné transversalement et les basidiospores sont alors émises latéralement par chacune des loges, comme chez les Urédinales, ou non cloisonné, émettant un bouquet de basidiospores portées par des stérigmates apicaux, comme chez les Holobasidiomycètes. Sur la base de ce critère distinctif, on répartit habituellement les Ustilaginales en deux familles distinctes, respectivement les Ustilaginacées et les Tillétiacées. Il a même été proposé de démanteler l'ordre des Ustilaginales pour rapprocher les Ustilaginacées des Urédinales et des Phragmobasidiomycètes et placer, au contraire, les Tillétiacées au voisinage des Holobasidiomycètes. Toutefois, si l'on tient compte à la fois de la relative homogénéité du comportement biologique des Ustilaginales et de la variabilité de leurs promycéliums, dont la morphologie n'est pas toujours aussi figée que les présentations synthétiques tendent à le suggérer, il apparaît excessif d'opposer aussi radicalement les deux familles d'Ustilaginales.
Accédez à l'intégralité de nos articles
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Patrick JOLY : directeur de recherche au C.N.R.S.
Classification
Médias

Espèce de chanterelle
Steven Taylor/ The Image Bank/ Getty Images

Cèpe de Bordeaux
mongider/ Flickr ; CC- BY-SA 2.0

Coprin chevelu
A. Lucini/ De Agostini/ Getty Images
Autres références
-
CHAMPIGNONS
- Écrit par Jacques GUINBERTEAU , Patrick JOLY , Jacqueline NICOT et Jean Marc OLIVIER
- 10 963 mots
- 17 médias
 Lesbasidiomycètes sont des champignons supérieurs caractérisés par le fait que les spores postméiotiques ( basidiospores) ne sont pas produites à l'intérieur de la cellule mère comme chez les ascomycètes, mais à l'extrémité d'un diverticule (stérigmate) émis par la cellule mère ( baside). Celle-ci,...
Lesbasidiomycètes sont des champignons supérieurs caractérisés par le fait que les spores postméiotiques ( basidiospores) ne sont pas produites à l'intérieur de la cellule mère comme chez les ascomycètes, mais à l'extrémité d'un diverticule (stérigmate) émis par la cellule mère ( baside). Celle-ci,... -
GAMÈTES
- Écrit par Michel FAVRE-DUCHARTRE et Jacques TESTART
- 4 174 mots
- 5 médias
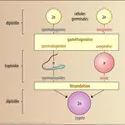 – chez les Basidiomycètes, la mise en commun des protoplasmes gamétiques est assurée par la conjonction des filaments mycéliens (somatogamie), pas même renflés en ampoules ;
– chez les Basidiomycètes, la mise en commun des protoplasmes gamétiques est assurée par la conjonction des filaments mycéliens (somatogamie), pas même renflés en ampoules ; -
LEVURES
- Écrit par Jacques BOIDIN , Encyclopædia Universalis , Jean-Bernard FIOL et Simone PONCET
- 3 438 mots
- 2 médias
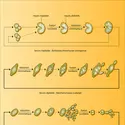 ...causes diverses – par exemple, maintien de l'état haploïde –, ne forment pas d'ascospores, pour une autre part de Champignons supérieurs, Euascomycètes et Basidiomycètes ayant acquis, avec le bourgeonnement, des thalles dissociables et une vie plus ou moins franchement unicellulaire. Une similitude des...
...causes diverses – par exemple, maintien de l'état haploïde –, ne forment pas d'ascospores, pour une autre part de Champignons supérieurs, Euascomycètes et Basidiomycètes ayant acquis, avec le bourgeonnement, des thalles dissociables et une vie plus ou moins franchement unicellulaire. Une similitude des... -
MICROBIOLOGIE
- Écrit par Corinne DOREL , Philippe LEJEUNE et Jean-Michel PANOFF
- 3 881 mots
- 9 médias
 ...sexuée ou les deux. Enfin, ils sont présents dans les quatre embranchements connus du règne des champignons, les phycomycètes, les ascomycètes, les basidiomycètes et les deutéromycètes. De manière tout à fait caractéristique, les phycomycètes (par exemple, Rhizopus stolonifer, moisissure commune...
...sexuée ou les deux. Enfin, ils sont présents dans les quatre embranchements connus du règne des champignons, les phycomycètes, les ascomycètes, les basidiomycètes et les deutéromycètes. De manière tout à fait caractéristique, les phycomycètes (par exemple, Rhizopus stolonifer, moisissure commune...
Voir aussi
- BASIDES
- AURICULARIALES
- APHYLLOPHORALES
- AGARICALES
- AMANITES
- BOLET
- HYPHES
- HÉTÉROTHALLISME
- DICARYONS
- URÉDINALES
- ROUILLE, phytopathologie
- TRÉMELLALES
- RUSSULALES
- SAPROPHYTISME
- MYCÉLIUM
- DACRYMYCÉTALES
- EXOBASIDIALES
- HOLOBASIDIOMYCÈTES
- PHRAGMOBASIDIOMYCÈTES
- PILEUS, botanique
- PLUTÉALES
- TRICHOLOMATALES
- TULASNELLALES
- TÉLIOBASIDIOMYCÈTES
- BASIDIOSPORES
- ANGIOCARPIE
- ÉCIDIE & ÉCIDIOSPORE
- URÉDOSPORE
- TÉLEUTOSPORE
