CELLULE L'organisation
Article modifié le
Tous les organismes sont constitués d'une ou plusieurs cellules, unités élémentaires dont la taille est de quelques dizaines de micromètres (1 μm = 10—3 mm). Bien inférieure au pouvoir séparateur de notre œil, cette petite taille des cellules explique qu'elles n'aient pu être observées avant l'invention du microscope au xviie siècle. Les bactéries, les protozoaires comme l'amibe ou la paramécie sont des organismes unicellulaires, les plantes (à l'exception de certains champignons comme les levures ou de certaines algues comme les diatomées) et les animaux sont des organismes pluricellulaires.
L'essentiel de ce que nous savons aujourd'hui sur la cellule a été acquis depuis 1955 ; à cette époque ont été mises au point les méthodes permettant l'observation au microscope électronique de coupes minces de cellules dont l'épaisseur est inférieure à 100 nanomètres (1 nm = 10—6 mm). C'est ainsi qu'ont été découverts les détails de l'anatomie cellulaire, détails que le microscope à lumière ne permet pas de voir et qui constituent l'ultrastructure de la cellule.

Simultanément, les techniques de fractionnement ont permis, à partir de populations cellulaires (cellules en culture, fragments d'organes), d'isoler des fractions ne contenant qu'une catégorie d' organites ; dans ces conditions on peut analyser la composition chimique des organites et même dans certains cas étudier leur fonctionnement hors de la cellule. À ce développement rapide des connaissances ont collaboré les cytologistes, les biochimistes, les biologistes moléculaires, les généticiens, les physiologistes, si bien que la biologie cellulaire est devenue un des thèmes les plus importants de la biologie fondamentale.
Malgré la diversité des êtres vivants, les cellules ont des caractères anatomiques, biochimiques et fonctionnels communs. Les pièces qui les composent sont très semblables, comme le sont également les molécules porteuses de leur information ou celles qui interviennent dans les conversions d'énergie et permettent à la cellule d'effectuer des travaux variés. Pour leur croissance, pour leur reproduction par division, pour la régulation de leurs activités, les cellules mettent en jeu les mêmes types de mécanismes. Cette unité du monde vivant à l'échelle cellulaire est l'héritage d'une longue évolution commencée il y a 4,5 milliards d'années avec la formation de la Terre, évolution qui a conduit à l'apparition des premiers organismes unicellulaires il y a 3,5 milliards d'années et qui s'est poursuivie jusqu'à maintenant.
L'anatomie cellulaire
Quel que soit l'organisme auquel elles appartiennent, les cellules sont formées de pièces qui remplissent des fonctions spécialisées tout comme les organes d'un être complexe, c'est pourquoi on les appelle organites cellulaires. L'étude anatomique des cellules faite à l'échelle la plus fine, grâce en particulier au microscope électronique, révèle que les organites ont une structure caractéristique qui est en quelque sorte leur « photo d'identité ». Les analyses biochimiques et biophysiques révèlent la nature des molécules qui composent les organites et, pour nombre d'entre eux, on connaît la disposition spatiale de leurs molécules, c'est-à-dire leur architecture moléculaire.

Noyau cellulaire
Planeta Actimedia S.A.© Encyclopædia Universalis France pour la version française.
Au centre de la cellule existe une région où est rassemblée sa « mémoire » opérationnelle : c'est le noyau qui est entouré par le cytoplasme. Chez les bactéries et les cyanobactéries, aucune compartimentation ne sépare le matériel nucléaire (nucléoïde) et le cytoplasme : ces organismes sont les procaryotes. Chez les animaux et les végétaux, une enveloppe nucléaire sépare le noyau du cytoplasme ; ces organismes dont les cellules ont un noyau individualisé sont les eucaryotes.

Cytoplasme cellulaire
Planeta Actimedia S.A.© Encyclopædia Universalis France pour la version française.

Cellule eucaryote
Planeta Actimedia S.A.© Encyclopædia Universalis France pour la version française.
Cellules procaryotes : bactéries et cyanobactéries

Cellules procaryotes : anatomie
Encyclopædia Universalis France
Une bactérie comme le colibacille ( Escherichia coli) est une cellule qui a la forme d'un bâtonnet cylindrique aux extrémités arrondies ; elle mesure 2,5 μm de long et 0,8 μm de diamètre (fig. 1a).
Cette cellule est enfermée à l'intérieur d'une enveloppe rigide ou paroi formée par l'association complexe de polypeptides, de polysaccharides et de lipides qui ont été synthétisés par le microbe lui-même. La paroi de 10 nm impose sa forme à la bactérie, elle représente donc un renforcement extérieur à la cellule, en dessous duquel la périphérie cellulaire est délimitée par une couche continue de 7,5 nm d'épaisseur qui est sa membrane plasmique. Cette membrane sépare l'intérieur de la cellule du milieu extracellulaire : c'est la surface cellulaire, constituée d'un assemblage de protéines et de lipides, qui contrôle les échanges entre la bactérie et le milieu où elle vit, que ce soit le contenu de l'intestin humain ou des milieux d'eau douce variés. Quant au cytoplasme, c'est un milieu visqueux riche en protéines, le cytosol, dans lequel baignent plus de 10 000 particules de 25 nm de diamètre, les ribosomes. Formées d'acides ribonucléiques, les ARN ribosomiques (ARNr), associés à des protéines, les ribosomes sont les organites cytoplasmiques qui synthétisent les chaînes polypeptidiques des protéines.
Le nucléoïde qui occupe la région centrale de la cellule est un filament de 2 nm de diamètre et de 1,2 mm de long ; refermé sur lui-même et reployé de nombreuses fois (fig. 1b). C'est une molécule circulaire d'acide désoxyribonucléique ( ADN) qui forme le chromosome bactérien parce qu'il est porteur de l'identité de l'espèce, codée dans la structure de la molécule. Celle-ci est donc qualifiée d'informationnelle.
Le chromosome n'est pas le seul organite porteur d'information. Il existe en effet dans le cytoplasme colibacillaire de petites molécules d'ADN circulaire de quelques microns de circonférence, que l'on appelle plasmides. Les plasmides étant en général tous identiques, l'information extrachromosomique qu'ils portent est donc présente en plusieurs exemplaires dans la cellule, alors que l'information chromosomique n'est représentée qu'à un seul exemplaire en dehors des périodes de réplication de cette molécule, qui permettront à la cellule de colibacille de pouvoir se dédoubler, autrement dit se reproduire.
Pour croître et se multiplier, le colibacille doit puiser dans le milieu qui l'environne des sels minéraux et des molécules organiques comme le glucose ; ces molécules lui servent à la fois de matériaux de construction et de source d'énergie. De tels organismes qui ont besoin de molécules organiques comme source de carbone sont dits hétérotrophes. D'autres bactéries synthétisent leurs molécules organiques à partir du gaz carbonique et de l'azote, l'énergie nécessaire à ces synthèses étant fournie par la lumière selon un mécanisme complexe qui est la photosynthèse. Ces organismes qui n'ont pas besoin de molécules organiques comme source de carbone sont dits autotrophes. Les bactéries photosynthétiques possèdent des organites cytoplasmiques originaux, responsables de leur caractère autotrophe : il s'agit de membranes de 6 nm d'épaisseur qui délimitent des cavités closes : les thylakoïdes. Selon les espèces, les thylakoïdes forment des vésicules de 50 nm de diamètre ou des sacs aplatis. La membrane des thylakoïdes est constituée de lipides et de protéines différents de ceux de la membrane plasmique et, de plus, elle renferme des pigments dont une variété de chlorophylle, qui captent l'énergie lumineuse ; ce type de membrane isole du cytosol un compartiment qui est l'espace intrathylakoïde.
Une cyanobactérie comme l'oscillaire (fig. 1c) est un organisme photosynthétique pluricellulaire qui vit dans l'eau des mares ; c'est un filament cylindrique de 4 μm de diamètre et de plusieurs centaines de microns de longueur qui est constitué de cellules identiques placées bout à bout. Chaque cellule, limitée par une membrane plasmique, est enfermée dans une paroi comparable à celle du colibacille et, en outre, la surface latérale de la paroi est recouverte extérieurement d'une couche de mucilage de 50 nm d'épaisseur. Ce mucilage est riche en polysaccharides qui, comme les constituants de la paroi, sont synthétisés par la cellule. Des thylakoïdes à disposition concentrique sont situés à la périphérie du cytoplasme. Leur membrane renferme de la chlorophylle et d'autres pigments qui donnent aux cellules une couleur bleue, d'où le nom d'algue bleue qui était donné à ces organismes (aujourd'hui le terme algue est réservé à des végétaux eucaryotes).
Les ribosomes des cellules d'oscillaire sont situés entre les thylakoïdes périphériques et le nucléoïde central, nucléoïde formé d'un chromosome sans doute circulaire et de 3 μm de circonférence. Le cytosol renferme des plasmides et des inclusions qui sont des réserves intracellulaires de nature chimique variée : gouttelettes de lipides, corps protéiques.
Cellules eucaryotes animales et végétales
Cellule animale
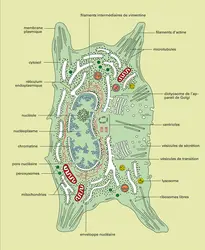
Cellule animale : anatomie
Encyclopædia Universalis France
Les fibroblastes humains sont les cellules spécifiques du tissu conjonctif comme le derme de la peau, le chorion du tube digestif, les tendons ou la paroi des veines et des artères ; ce sont des cellules isolées qui baignent dans le milieu extracellulaire contenant de nombreuses fibres protéiques de collagène et d'élastine ainsi que des macromolécules complexes – les protéoglycanes – formées de chaînes polypeptidiques auxquelles sont attachées de longues chaînes polysaccharidiques sulfatées. Toutes ces molécules qui sont synthétisées par les fibroblastes donnent au tissu conjonctif ses propriétés mécaniques d'élasticité et de résistance à l'extension. Un fibroblaste (fig. 2) a la forme d'un fuseau en périphérie duquel s'étendent divers prolongements. Le corps cellulaire mesure 20 μm de long sur 5 μm de diamètre environ, les prolongements de longueur variée ont un diamètre souvent inférieur au micromètre. Le noyau fusiforme occupe la région centrale et comme chez les procaryotes, le corps cellulaire est limité extérieurement par une membrane plasmique de 7,5 nm d'épaisseur.
L'anatomie de cette cellule est tout d'abord caractérisée par la présence dans son cytoplasme de différents types de membranes qui isolent du cytosol des compartiments clos, représentant autant d'organites originaux.
En premier lieu, les membranes du réticulum endoplasmique (6 nm d'épaisseur) délimitent un ensemble de cavités ou citernes qui communiquent entre elles et forment un réseau spongieux à l'intérieur du cytoplasme (d'où le nom qui a été donné à cet organite). Une portion du réticulum entoure le noyau, c'est l'enveloppe nucléaire qui sera étudiée en détail plus loin. Elle marque la frontière entre le noyau et le cytoplasme ; cette frontière n'est pas continue car elle est fenestrée de place en place par des perforations de 50 nm de diamètre qui sont les pores nucléaires.
De nombreux ribosomes sont attachés aux membranes de réticulum (fig. 3) de cette cellule et leur donnent un aspect grenu, d'où le nom de réticulum endoplasmique rugueux attribué à ce type de réticulum (on l'appelle aussi ergastoplasme). Les chaînes polypeptidiques qui sont synthétisées par ces ribosomes sont transférées dans les cavités du réticulum ou insérées dans les membranes. Les portions de réticulum dont les membranes sont dépourvues de ribosomes constituent le réticulum endoplasmique lisse, type de réticulum qui est peu abondant dans le fibroblaste.
Les membranes de l'appareil de Golgi (7 nm d'épaisseur et toujours dépourvues de ribosomes) isolent des cavités dont l'arrangement est beaucoup plus ordonné que le réseau polymorphe du réticulum endoplasmique. Il s'agit de disques aplatis ou saccules qui sont empilés les uns sur les autres ; chaque pile est un dictyosome et compte 3 à 4 saccules de 1 μm de diamètre environ ; au nombre d'une dizaine, les dictyosomes sont groupés près du noyau.
De part et d'autre des dictyosomes existent deux populations de vésicules de 20 nm de diamètre : des vésicules de transition situées entre les dictyosomes et des citernes de réticulum, des vésicules de sécrétion situées au voisinage de l'autre face des dictyosomes. Ces deux populations de vésicules témoignent de la structure dynamique des dictyosomes : de nouveaux saccules se forment à la face externe par fusion de vésicules de transition qui ont bourgeonné à partir du réticulum, en même temps que des saccules disparaissent à la face opposée en se fragmentant en vésicules de sécrétion.
Les membranes du réticulum endoplasmique contiennent des enzymes dont certaines catalysent la synthèse de certains lipides membranaires, d'autres la liaison covalente de sucres à des molécules variées, c'est-à-dire qu'elles catalysent des réactions du glycosylation, d'autres enfin, qui inactivent des substances toxiques pour la cellule et interviennent donc dans les détoxifications (cf. détoxification). Les membranes de l'appareil de Golgi interviennent elles aussi dans des réactions de glycosylation et de plus dans la sulfatation de certaines molécules auxquelles sont ainsi liés de façon covalente des groupements sulfate.
Les cavités du réticulum forment des compartiments où s'accumulent des molécules destinées à être exportées dans le milieu extracellulaire ou à être stockées dans d'autres compartiments intracytoplasmiques. Après avoir été synthétisées par les ribosomes du réticulum rugueux et transférées dans ses cavités, les chaînes polypeptidiques du collagène, de l'élastine (cf. collagène) et des protéoglycanes caractéristiques de la matrice du tissu conjonctif cheminent dans ce réseau cavitaire jusqu'aux régions situées en regard des dictyosomes ; là elles quittent le réticulum dans les vésicules de transition, elles transitent dans un saccule de la face externe d'un dictyosome jusqu'à sa face interne où elles sont alors emballées dans des vésicules de sécrétion. Au cours de ce cheminement intracellulaire les chaînes polypeptidiques sont toujours séparées du cytosol par une membrane (d'abord du réticulum puis de l'appareil de Golgi), et de plus elles sont modifiées chimiquement, les réactions étant catalysées par des enzymes situées dans les cavités que limitent les membranes de ces organites. Les vésicules de sécrétion sont emportées vers la périphérie du fibroblaste et leur contenu est alors déchargé dans le milieu extracellulaire par fusion de leur membrane avec la membrane plasmique du fibroblaste selon un processus appelé exocytose. Dans le milieu extracellulaire, les molécules de collagène ou d'élastine s'associent entre elles en formant les fibres du tissu conjonctif dans lequel le fibroblaste est placé.
Dans le cytoplasme du fibroblaste existent également d'autres organites limités par des membranes et qui sont en fait des productions du réticulum endoplasmique ou de l'appareil de Golgi. Il s'agit tout d'abord de vésicules de 0,1 μm de diamètre, limitées par une membrane de 6 nm d'épaisseur qui isole une matrice ; dispersées dans le cytosol, ces vésicules renferment des enzymes qui catalysent la production et la décomposition de l'eau oxygénée, c'est-à-dire le peroxyde d'hydrogène ; ces organites sont les peroxysomes. D'autres vésicules de diamètre varié (0,2 μm à 2 ou 3 μm) limitées par une membrane de 7 nm d'épaisseur contiennent un mélange d' hydrolases, enzymes qui catalysent à pH acide le clivage de molécules variées avec intervention de molécules d'eau ; ces organites sont les lysosomes qui permettent au fibroblaste de digérer les molécules qu'il capture dans le milieu extracellulaire.
Les peroxysomes proviennent de bourgeons qui se détachent du réticulum endoplasmique ; les enzymes de leur matrice sont synthétisées pour une part au niveau de ribosomes liés aux membranes du réticulum, pour une autre part au niveau de ribosomes libres dans le cytosol et transférées après leur synthèse dans la matrice. Les lysosomes sont bourgeonnés par les bords des saccules golgiens, leurs hydrolases étant synthétisées par des ribosomes du réticulum rugueux ; ces enzymes empruntent au départ la même voie de cheminement que les chaînes polypeptidiques destinées à l'exportation, mais elles sont ensuite aiguillées différemment.
Les mitochondries représentent un dernier type d'organite cytoplasmique à structure membranaire. Au nombre d'une centaine dispersées dans le cytosol, les mitochondries du fibroblaste ont la forme de bâtonnets cylindriques aux extrémités arrondies dont le diamètre est de 0,5 μm et la longueur de 2 à quelques micromètres. Chaque mitochondrie est séparée du cytosol par une membrane de 6 nm d'épaisseur qui est la membrane mitochondriale externe ; cette membrane est doublée intérieurement d'une membrane mitochondriale interne également de 6 nm d'épaisseur et qui forme des replis appelés crêtes mitochondriales. Les deux membranes mitochondriales isolent deux compartiments : l'un situé entre les deux membranes externe et interne qui est l'espace intermembranaire, un autre limité par la membrane interne qui est la matrice mitochondriale. Les mitochondries régénèrent le fournisseur universel d'énergie pour la cellule : l'adénosine triphosphate ou ATP (cf. bioénergétique). L'hydrolyse de ce nucléotide donne de l'adénosine diphosphate ou ADP, de l'acide phosphorique et de l'énergie, énergie qui est utilisée pour des travaux cellulaires de nature variée : synthèse de molécules, production du mouvement, transport d'ions d'un compartiment à l'autre par exemple. La régénération des molécules d'ATP se fait par phosphorylation de l'ADP en ATP ; cette phosphorylation nécessite un apport énergétique qui est fourni cette fois par l'oxydation de nutriments organiques, surtout glucidiques ou lipidiques, qui représentent en quelque sorte des « combustibles » bioénergétiques. Les oxydations se déroulent principalement dans la matrice mitochondriale qui contient de nombreuses enzymes catalysant ces réactions ; le couplage énergétique de ces réactions d'oxydation à la phosphorylation de l'ADP est la phosphorylation oxydative : elle se fait au niveau de la membrane mitochondriale interne et met en jeu des protéines spécifiques de cette membrane.
Outre les organites à membranes et les nombreux ribosomes dispersés dans le cytosol ou associés aux membranes du réticulum endoplasmique, existent dans le cytoplasme des structures filamenteuses qui interviennent dans le maintien de la forme du fibroblaste et dans les mouvements de cette cellule ; ces structures forment un cytosquelette intracellulaire comprenant trois types d'organites : des microtubules, des filaments d'actine et des filaments intermédiaires.
Les microtubules sont des tubes cylindrique de 25 nm de diamètre et dont la paroi mesure 5 nm d'épaisseur ; les uns sont courts (0,3 μm) et associés par groupes de 3 comme les génératrices d'un cylindre : ce sont les deux centrioles qui sont constitués chacun de 9 groupes de 3 microtubules ; les autres sont longs (de 3 μm à 10 μm) et isolés. En disposition rayonnante à partir de la région péricentriolaire, ces filaments s'étendent jusque dans les prolongements cytoplasmiques du fibroblaste. La paroi des microtubules est constituée de protéines globulaires appelées tubulines.
Les filaments d'actine, d'un diamètre de 5 nm et de plusieurs microns de long sont pour la plupart groupés en faisceaux ou câbles présents dans le corps cellulaire et aussi dans les prolongements périphériques où leur orientation est parallèle à celle des microtubules. L'actine qui compose ces filaments est une protéine globulaire.
Les filaments intermédiaires ont un diamètre de 10 nm, diamètre compris entre celui des microtubules et celui des filaments d'actine. Dans le fibroblaste ces filaments sont formés de vimentine, qui est une protéine fibreuse ; ils forment un lacis situé principalement autour du noyau qu'ils maintiennent au centre de la cellule.

Chromosomes humains
Dan McCoy-Rainbow/Age Fotostock/ Imagestate
L'enveloppe nucléaire qui délimite le noyau sépare deux milieux riches en protéines : le cytosol et le nucléoplasme. Dans le nucléoplasme baignent des filaments de 10 nm de diamètre appelés nucléofilaments qui, dans les fibroblastes en cours de reproduction, deviendront les chromosomes, organites portant l'essentiel de l'information cellulaire car ils contiennent de l'ADN. Chez l'homme, les chromosomes sont au nombre de 46. Chaque chromosome est constitué d'une molécule d'ADN linéaire à laquelle sont associées des protéines. Les unes compactent l'ADN du nucléofilament : ce sont les histones (en absence d'histones, les 46 molécules d'ADN ont une longueur totale de 2 m environ). Les autres ont une activité catalytique dans la synthèse de molécules d' ARN ou la synthèse de nouvelles molécules d'ADN : ce sont des enzymes.
Les nucléofilaments sont plus ou moins condensés et enchevêtrés dans le noyau et forment ainsi des masses fibreuses visibles au microscope à lumière. La colorabilité de ce matériau par les colorants basiques (vert de méthyle par exemple) lui ont valu son nom de chromatine ; selon son état de compaction on distingue la chromatine condensée et la chromatine diffuse.
Le noyau renferme de nombreuses particules ribonucléoprotéiques ou particules RNP dont le diamètre est compris entre 5 nm et 20 nm. L'ARN des particules RNP est synthétisé au niveau des chromosomes ; leurs protéines viennent du cytoplasme où elles sont synthétisées par les ribosomes libres. Les particules RNP qui s'assemblent dans le noyau sont ensuite exportées dans le cytoplasme en passant à travers les pores de l'enveloppe nucléaire. Il existe une catégorie de particules RNP qui sont produites en grande quantité dans le noyau, ce sont les préribosomes, particules précurseurs des ribosomes cytoplasmiques. Les préribosomes sont stockés transitoirement dans le noyau avant leur exportation, ce stock forme une masse particulaire sphérique de 1 μm de diamètre : le nucléole.
En plus des chromosomes nucléaires, le fibroblaste possède d'autres structures porteuses d' information pour la synthèse protéique et la réplication de l'organite qui les renferme. Ce sont de petites molécules d'ADN circulaires de 5 μm de circonférence : les molécules d'ADN mitochondrial ou ADNmt. Dans sa matrice chaque mitochondrie renferme 3 à 5 molécules d'ADNmt et également des ribosomes différents de ceux du cytoplasme ; ces ribosomes ou mitoribosomes synthétisent certaines des chaînes polypeptidiques de la membrane mitochondriale interne.
Cellule végétale
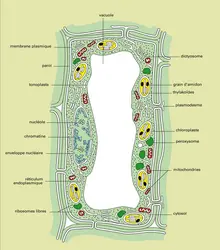
Cellule végétale : anatomie
Encyclopædia Universalis France
Une cellule du tissu vert friable que renferme une feuille d'épinard (parenchyme palissadique) d'une feuille d'épinard est de forme prismatique et mesure 60 μm de haut et 20 μm de large. De forme prismatique (fig. 4), elle est délimitée par une paroi rigide de 0,1 μm d'épaisseur, riche en polysaccharides. C'est un cytosquelette extracellulaire dont les constituants sont synthétisés par la cellule elle-même ; elle est formée de fibrilles de cellulose disposées parallèlement à la surface cellulaire et dont les diamètres sont compris entre 3,5 μm et 10 μm. Ces fibrilles sont enrobées dans une matrice formée d'hémicellulose et de substances pectiques. Cette paroi est tapissée par une membrane plasmique plaquée contre sa face interne. Au centre du cytoplasme se trouve un lysosome volumineux et très hydraté qui est la vacuole. Les autres organites cellulaires sont situés en périphérie, entre la membrane de la vacuole (appelée tonoplaste) et la membrane plasmique ; réticulum endoplasmique rugueux et lisse, ribosomes libres ou attachés aux membranes du réticulum, dictyosomes de l'appareil de Golgi, mitochondries, peroxysomes, noyau avec son enveloppe nucléaire et contenant de la chromatine ainsi que un ou deux nucléoles sont donc présents comme dans le fibroblaste.
Le cytosquelette intracellulaire comprend des microtubules et des filaments d'actine localisés principalement sous la surface cellulaire. Comme la plupart des cellules végétales, cette cellule de parenchyme palissadique ne possède pas de centrioles mais elle renferme des organites originaux qui contiennent de la chlorophylle : les chloroplastes, au nombre d'une centaine (fig. 5).
Chaque chloroplaste est un disque lenticulaire de 5 ou 6 μm de diamètre et de 2 μm d'épaisseur dans sa partie centrale ; il est limité par une enveloppe formée de deux membranes concentriques. L'enveloppe isole un compartiment qui est le stroma du chloroplaste, stroma contenant des thylakoïdes dont la membrane a 7 nm d'épaisseur. Les thylakoïdes sont orientés parallèlement au grand axe du chloroplaste et certains sont empilés ; chaque pile est un « granum » et on compte 40 à 60 granums par chloroplaste. La membrane des thylakoïdes contient de la chlorophylle et des pigments qui donnent aux feuillets leur couleur verte. Grâce à leurs chloroplastes, les cellules du parenchyme réalisent la photosynthèse ; l'énergie lumineuse captée au niveau des thylakoïdes permet la réduction du gaz carbonique et d'autres composés minéraux en molécules organiques glucidiques, par exemple des molécules d'amidon qui sont stockées temporairement sous forme de grains d'amidon dans le stroma.
Le stroma contient des molécules d'ADN circulaire de 50 μm de circonférence et qui sont au nombre d'une vingtaine par chloroplaste : ce sont les molécules d'ADN chloroplastique ou ADNct ; de plus le stroma renferme des plastoribosomes différents de ceux du cytoplasme et des mitoribosomes ; les plastoribosomes synthétisent des chaînes polypeptidiques de la membrane des thylakoïdes et certaines enzymes du stroma.
Les cellules du parenchyme communiquent entre elles par des ponts cytoplasmiques qui traversent leur paroi : les plasmodesmes au nombre de plusieurs milliers par cellule ; certains plasmodesmes renferment un tube de réticulum endoplasmique lisse qui passe d'une cellule à sa voisine.

Arrangement cellulaire
DeAgostini/ Getty Images
Au total, sur le plan structural, les cellules ne sont que des variations sur deux grands thèmes : le thème procaryote et le thème eucaryote ; ces deux thèmes ont d'ailleurs des points communs puisque la membrane plasmique, les chromosomes, les ribosomes sont des organites qui existent dans toutes les cellules et que les thylakoïdes sont caractéristiques des cellules photosynthétiques possédant de la chlorophylle, qu'elles soient procaryotes ou eucaryotes.
Accédez à l'intégralité de nos articles
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Pierre FAVARD : professeur à l'université de Paris-VI-Pierre-et-Marie-Curie
Classification
Médias
Noyau cellulaire
Planeta Actimedia S.A.© Encyclopædia Universalis France pour la version française.
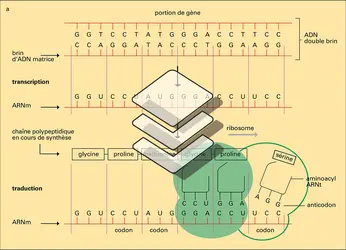
Information cellulaire
Encyclopædia Universalis France
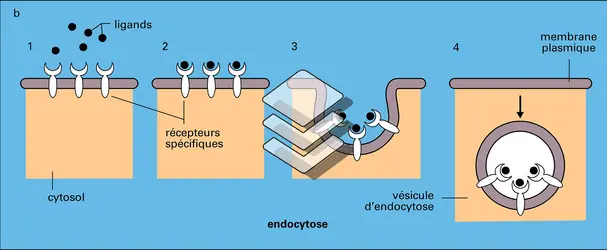
Échanges avec le milieu extracellulaire
Encyclopædia Universalis France
Autres références
-
CELLULE, notion de
- Écrit par Didier LAVERGNE
- 1 315 mots
- 5 médias

Le terme cellule (cell en anglais) apparaît en 1665, sous la plume du physicien anglais Robert Hooke, pour désigner les logettes que l'on voit au microscope dans un fragment végétal inerte, le liège. En 1824, le biologiste français Henri Dutrochet, qui s'intéresse aux forces agissant au sein des...
-
COMMUNICATION CELLULAIRE
- Écrit par Yves COMBARNOUS
- 6 593 mots
- 7 médias
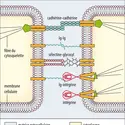
-
MITOSE
- Écrit par Nina FAVARD
- 6 522 mots
- 5 médias
L'aphorisme omnis cellula e cellula (« toute cellule est issue d'une autre cellule ») a permis aux biologistes de comprendre que, au cours des générations cellulaires successives, la transmission de l'information génétique obéissait à des mécanismes d'une grande précision....
-
ADN (acide désoxyribonucléique) ou DNA (deoxyribonucleic acid)
- Écrit par Michel DUGUET , Encyclopædia Universalis , David MONCHAUD et Michel MORANGE
- 10 074 mots
- 10 médias
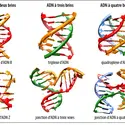 D'où le formidable intérêt pour lacellule de posséder un ADN circulaire fermé : le nombre de liens topologiques L entre les deux brins y est différent de zéro. Si, donc, la cellule dispose d'un moyen de réguler la valeur de L, elle pourra du même coup (L = T + W) fixer la valeur...
D'où le formidable intérêt pour lacellule de posséder un ADN circulaire fermé : le nombre de liens topologiques L entre les deux brins y est différent de zéro. Si, donc, la cellule dispose d'un moyen de réguler la valeur de L, elle pourra du même coup (L = T + W) fixer la valeur... -
ANIMAUX MODES DE REPRODUCTION DES
- Écrit par Catherine ZILLER
- 4 448 mots
- 4 médias
 Lorsqu'unanimal pluricellulaire atteint l'état adulte, la majorité de ses cellules sont spécialisées. Elles sont physiologiquement et morphologiquement différenciées ; elles constituent les tissus et les organes dont cet animal a besoin pour survivre : elles forment ce que l'on appelle le ...
Lorsqu'unanimal pluricellulaire atteint l'état adulte, la majorité de ses cellules sont spécialisées. Elles sont physiologiquement et morphologiquement différenciées ; elles constituent les tissus et les organes dont cet animal a besoin pour survivre : elles forment ce que l'on appelle le ... -
ANIMAUX MODÈLES, biologie
- Écrit par Gabriel GACHELIN et Emmanuelle SIDOT
- 9 550 mots
- 8 médias
 Depuis de nombreuses décennies – la première mise en culture au long terme decellules animales remonte à 1912 –, les scientifiques utilisent en routine des lignées cellulaires, cellules dites « immortelles », leur culture – division cellulaire in vitro – pouvant s’effectuer indéfiniment tant qu’elles...
Depuis de nombreuses décennies – la première mise en culture au long terme decellules animales remonte à 1912 –, les scientifiques utilisent en routine des lignées cellulaires, cellules dites « immortelles », leur culture – division cellulaire in vitro – pouvant s’effectuer indéfiniment tant qu’elles... - Afficher les 139 références
Voir aussi
- ENZYMES
- FIBROBLASTE
- TRANSPORTS MEMBRANAIRES
- CATALYSE ENZYMATIQUE
- CYTOSOL
- ESCHERICHIA COLI ou COLIBACILLE
- CYTOPLASME
- RÉTICULUM ENDOPLASMIQUE
- GOLGI APPAREIL DE
- DICTYOSOMES
- PINOCYTOSE
- NOYAU CELLULAIRE
- NUCLÉOTIDES
- CODON, biologie moléculaire
- RÉPLICATION, biologie moléculaire
- TRANSCRIPTION, biologie moléculaire
- TRADUCTION, biologie moléculaire
- STROMA
- THYLAKOÏDES
- INFECTION
- MICROTUBULE, biologie
- MICROFILAMENT, biologie
- FILAMENT INTERMÉDIAIRE, biologie
- PECTO-CELLULOSIQUE PAROI
- MÉTABOLISME CELLULAIRE
- POLYPEPTIDIQUE CHAÎNE
- PHOSPHORYLATION
- MÉDIATEURS BIOCHIMIQUES
- NICOTINAMIDE ADÉNINE DINUCLÉOTIDE PHOSPHATE (NADP)
- HYDROLASES
- PEPTIDIQUES HORMONES
- PHOSPHORYLATION OXYDATIVE
- ABSORPTION INTESTINALE
- PROTÉINES MEMBRANAIRES
- BIOLOGIE MOLÉCULAIRE
- MEMBRANES BIOLOGIQUES
- ORGANITES CELLULAIRES
- HISTONES
- AUTOASSEMBLAGE
- DIVISION CELLULAIRE
- CHROMOSOMES
- CHROMATINE
- BIOSYNTHÈSES
- JONCTIONS INTERCELLULAIRES
- PROCARYOTES ou PROTOCARYOTES
- INFORMATION GÉNÉTIQUE
- BIOÉNERGÉTIQUE
- CHAÎNE RESPIRATOIRE
- CAPSIDE
- RECOMBINAISON GÉNÉTIQUE
- PEROXYSOME
- PERMÉABILITÉ SÉLECTIVE
- DIFFUSION TRANSMEMBRANAIRE, biologie
- ATPASES (adénosine-tri-phosphatases)
- CYTOLOGIE ou BIOLOGIE CELLULAIRE
- TRANSPORT D'ÉLECTRONS, bioénergétique
- FORCE PROTOMOTRICE
- COUPLAGE, biochimie
- NEUROMÉDIATEURS ou NEUROTRANSMETTEURS
- GLYCOSYLATION
- SIGNAL, biologie
- ADÉNOSINE DIPHOSPHATE (ADP)
- CYTOLOGIE VÉGÉTALE
- TONOPLASTE, cytologie
- PROTONS GRADIENT DE
- CALCIUM & MÉTABOLISME CELLULAIRE
- GLYCOLIPIDES
- GLYCOPROTÉINES
- NUCLÉOTIDIQUE SÉQUENCE
- GÉNÉTIQUE MOLÉCULAIRE
- CALMODULINE
- EXOCYTOSE
- NUCLÉOFILAMENT
- PROTÉOGLYCANES
- PLASMODESME
- ARN RIBOSOMIQUE ou ARNr
- GRANUM
- PAROI CELLULAIRE
- PARENCHYME VÉGÉTAL
- PROTÉINES
- RÉCEPTEURS MEMBRANAIRES
- MUTAGENÈSE
- CYTOSQUELETTE
- KINASES
- VACUOLE
