CROISSANCE, biologie
Article modifié le
Biométrie : mensurations chez les organismes multicellulaires
Croissance absolue
On obtient la courbe de croissance absolue en mesurant la « taille » X à des intervalles suffisamment rapprochés, puis en la reportant sur un graphique en ordonnée, avec le temps en abscisse. De telles courbes peuvent être obtenues à partir d'un seul individu, quand elles ne perturbent pas la croissance et que celle-ci est rapide. On peut aussi mesurer de nombreux individus d'âges variés mais connus, par exemple chez l'homme, et en déduire la courbe moyenne de croissance pour l'espèce. L'emploi de ces moyennes n'est pas sans danger car, si elles font disparaître les petites fluctuations dues au hasard, elles risquent également de masquer des irrégularités de la courbe de croissance, accidents dont l'existence est constante et la signification importante, mais qui ne se produisent pas à un âge très précis.
Si l'on désigne par dX la croissance de X pendant un court intervalle de temps dt, le taux de croissance absolue est la limite vers laquelle tend le quotient dX/dt quand dt tend vers zéro, c'est-à-dire la valeur à l'instant t de la dérivée de X = f (t). Ce taux garde rarement une valeur constante et correspond alors à une partie linéaire de la courbe de croissance : les pousses de bambous en fournissent un exemple. Un paramètre plus intéressant est obtenu en divisant le taux défini précédemment par X, soit (1/X) . (dX/dt) ; il est souvent appelé taux de croissance relative, mais, en raison des confusions possibles avec les taux utilisés dans l'étude de la croissance relative, il vaut mieux parler de taux de croissance réduit, c'est-à-dire ramené à l'unité. Il est constant dans le cas d'une croissance exponentielle, ce qui se produit en général dans les premiers temps de la croissance, puis sa valeur décroît progressivement. Lorsque la courbe de croissance est tracée en coordonnées non plus arithmétiques mais semi-logarithmiques, c'est-à-dire lorsqu'on trace la courbe log X = f (t), ce taux de croissance réduit représente la valeur de la pente de la courbe au moment t.

Les courbes de croissance des organismes entiers montrent des aspects très variés. Dans les cas les plus simples, elles ont, comme pour une culture, l'allure d'une sigmoïde, plus ou moins symétrique. De nombreuses formules théoriques ont été proposées pour rendre compte de ces courbes ; établies à partir de considérations théoriques sur les mécanismes de la croissance, elles permettent de tester la valeur des hypothèses faites. La courbe logistique, de la forme 1/X = a (1 + be—ct), est obtenue par intégration de dX/dt = kX (F — X) ; cette formule, où F représente la taille finale, implique que le taux de croissance absolue est le produit de deux facteurs, l'un augmentant avec la taille (facteur auto-accélérateur), l'autre diminuant avec celle-ci (auto-inhibition). L'équation de Bertalanffy est déduite de dX/dt = aXm +bXn, le premier terme représentant les facteurs anaboliques et le second ceux du catabolisme ; si X est le poids, les paramètres m et n sont compris entre 2/3 et 1, ce qui est en conformité avec les relations existant entre métabolisme d'une part, surface et volume de l'autre. Ces deux équations permettent souvent d'arriver à un bon accord avec les courbes observées, mais il en est de même pour beaucoup d'autres dont la signification biologique est moins claire. Et la courbe observée est rarement obtenue avec une précision suffisante pour permettre de choisir entre les hypothèses.
Cette imprécision s'explique par deux raisons :
a) Il est exceptionnel qu'une seule loi de croissance reste valable pendant toute la vie.
Rat
Encyclopædia Universalis France
Le développement se compose presque toujours d'une succession de phases ayant chacune ses lois propres : le mode de vie et les conditions optimales de milieu changent souvent avec la phase, les différents organes se développant inégalement suivant les phases (fig. 2). L'examen attentif de courbes de croissance absolue du type sigmoïde révèle souvent qu'elles sont en réalité formées d'une succession d'arcs de courbe du type sigmoïde, exponentiel ou autre. La vie embryonnaire, la jeunesse, l'adolescence correspondent à de telles phases, ou même à plusieurs chacune.
b) Ces courbes ne sont valables qu'en milieu constant, car la vitesse de croissance à un moment donné dépend plus ou moins des conditions externes, par exemple de la qualité et de la quantité de nourriture : des carences alimentaires ralentissent ou même arrêtent la croissance. La température joue un rôle majeur chez les animaux à température variable ( poïkilothermes) comme cela a lieu, également, chez les végétaux. Quand la température interne diminue, tous les processus vitaux se ralentissent, comme pour des réactions chimiques banales, et cela est d'ailleurs un des points ayant conduit à la notion de temps biologique : mesurée en temps physique, son unité s'allonge quand la température baisse ; son emploi doit permettre d'obtenir des taux de croissance moins variables. Chez les arthropodes, l'existence des mues fournit une base objective à la mesure de ce temps biologique et permet d'en mettre la notion en pratique, par exemple en définissant le taux d'accroissement à la mue : ce rapport des tailles à deux stades successifs est relativement constant. La température n'est pas seule à agir sur l'unité de temps biologique. Il est banal de dire que le temps « passe plus vite » quand l'âge augmente, et ce ralentissement des fonctions vitales vient s'ajouter à celui qui est lié à l'augmentation de taille (courbe logistique). Cet allongement de l'unité de temps biologique avec la taille s'observe aussi quand on compare des espèces entre elles : des animaux semblables par leur morphologie et leur physiologie, mais de tailles adultes différentes, ont une vie d'autant plus courte et plus intense qu'ils sont plus petits.
L'intérêt des graphiques de croissance absolue est donc limité dans la pratique. Ils peuvent fournir, pour un milieu naturel défini, une approximation du temps nécessaire pour que soit atteinte une taille donnée, par exemple la taille minimale pour qu'un animal de pêche ou d'élevage soit commercialisable. En faisant varier expérimentalement certains facteurs du milieu, les courbes de croissance permettent également d'en évaluer l'influence.
Croissance relative
Loi d'allométrie
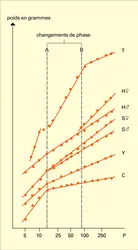
La croissance d'un organisme impliquant de nombreuses corrélations à tous les niveaux, on peut s'attendre qu'il existe, entre les croissances des diverses parties, des relations mathématiques plus ou moins indépendantes des facteurs externes. La plus fréquemment rencontrée de ces relations est la loi d'allométrie, de formule très simple : Y = BXa ; c'est une fonction puissance, et non une fonction exponentielle comme on l'écrit trop souvent. Elle se traduit en coordonnées logarithmiques par une droite y = ax + b, où x est le logarithme de la taille ou de tout autre caractère de référence et y celui du caractère qui lui est comparé ; le paramètre a porte le nom de taux d'allométrie, b celui d'indice d'origine. Pour des raisons de commodité, les deux paramètres a et b sont déterminés à partir d'un ensemble de points expérimentaux représentant non pas les divers états d'un individu au cours de sa vie, mais ceux, à un instant donné, du plus grand nombre possible d'individus de tailles variées ; cela permet d'ailleurs d'obtenir des courbes continues même si la croissance est intermittente.
Si a est égal à 1, il y a isométrie : Y croît proportionnellement à X ; l'isométrie est rare car, pour des raisons fonctionnelles, mécaniques et physiologiques, la forme doit changer avec la taille. Si a est supérieur à l'unité, il y a allométrie majorante : Y croît plus vite que X et il y a changement des proportions, de la forme. C'est le cas de nombreux variants sexuels qui se développent « brusquement » dans l'un des sexes, au moment de la puberté. L'un des premiers cas étudiés, et des plus spectaculaires, est celui des mâles d'Uca, le crabe violoniste (fig. 3). Les jeunes et les femelles ont de petites pinces, alors que l'une d'elles atteint une taille disproportionnée chez le mâle ; a étant égal à 1,6, le poids de la pince passe, pendant la puberté, de 10 à 40 p. 100 de celui du corps. Si a est inférieur à 1, il y a régression relative de Y, allométrie minorante, et, s'il est négatif, il y a régression en valeur absolue, ou énantiométrie.
Des relations d'allométrie ont été mises en évidence dans les domaines les plus divers : entre la taille d'une cellule et celle de son noyau (variations du rapport nucléoplasmatique) ; entre la taille, le poids et même le nombre d'organes, de membres, de soies, dans les groupes animaux les plus variés ; et même encore entre les teneurs en tel et tel constituant chimique de l'organisme. Les variations du taux d'allométrie d'un point à l'autre de l'organisme ne sont pas quelconques, car il y a corrélation.
Elles obéissent à des gradients, les taux diminuent progressivement de part et d'autre d'un ou plusieurs centres de croissance. Un gradient simple, fréquemment rencontré dans la croissance postembryonnaire, y compris celle de l'homme, est le gradient antéro-postérieur : selon l'axe du corps, le taux d'allométrie croît régulièrement à partir de la tête, ce qui permet de rattraper l'avance prise par la tête dès le début du développement embryonnaire, avance justifiée par l'importance des centres nerveux qu'elle abrite et le rôle de commande qu'elle joue très précocement. De nombreux cas de monstruosités s'expriment par des déformations minimes des gradients d'allométrie. L'existence des gradients, reflets des corrélations, explique aussi que la loi d'allométrie puisse s'appliquer de façon satisfaisante bien que la fonction puissance ne soit pas additive. Si Y est la somme de Y1 = B1Xa1 et de Y2 = B2Xa2, Y ne peut en théorie s'exprimer sous la forme BXa ; mais, si les coefficients des deux fonctions sont voisins, cette incompatibilité reste théorique, les écarts entre BXa et B1Xa1 + B2Xa2 étant très faibles, bien inférieurs à la dispersion des points expérimentaux.
La formule d'allométrie peut recevoir une interprétation physiologique très simple : elle signifie que les taux de croissance réduits de X et de Y sont, à un instant donné, proportionnels à la masse de nourriture disponible – et à elle seule –, selon un coefficient caractéristique de chacun d'eux, ainsi que de la phase et de l'espèce. De cette interprétation on tire une conclusion étonnante : il n'y a aucune relation de cause à effet entre la croissance des divers organes au cours d'une phase d'allométrie, les interactions ne peuvent jouer qu'au moment des changements de phase. Les exceptions apparentes à l'allométrie prouvent qu'il en est bien ainsi : l'allométrie ne peut maintenir l'ordre (limiter les variations) que s'il existe déjà ; elle ne peut le rétablir. Certains variants sexuels des Crustacés, par exemple, passent, sous l'influence de l' hormone sexuelle, par une phase d'allométrie fortement majorante lors de la puberté ; comme l'hormone déclenche cette puberté à une taille assez variable, l'indice d'origine b varie, et les points représentatifs d'un ensemble d'individus ne sont plus rassemblés en nuage étroit autour d'un segment d'allométrie, même quand chaque individu suit une loi d'allométrie (a est fixe). Au contraire, les points de la phase postpubérale s'alignent de nouveau autour d'un segment adulte parallèle à celui de la phase jeune. Comme l'hormone sexuelle reste présente dans le sang, il est clair que c'est un changement de propriétés des tissus qui a mis fin à la phase pubérale, et que ce changement est le résultat d'interactions moins bien connues mais plus précises que les mécanismes hormonaux. Si les tissus des variants sexuels n'étaient pas devenus insensibles à l'hormone et n'avaient pas retrouvé des taux d'allométrie plus normaux, leur croissance n'aurait pu se poursuivre sans aboutir à des animaux monstrueux, inviables.
Champ d'application
L'intérêt des courbes d'allométrie est beaucoup plus grand que celui des courbes de croissance absolue. À titre descriptif, elles permettent de présenter les faits sous une forme condensée, claire, objective, facilement transmissible. La croissance relative passe par la même série de phases que la croissance absolue, mais elle en permet plus facilement la mise en évidence. Chaque changement de phase se traduit par des modifications simultanées des lois d'allométrie pour un plus ou moins grand nombre de couples de caractères. Les phases sont représentées par des segments de droite séparés par des « points critiques », ou parfois des « discontinuités » qui correspondent en fait à des phases. Au long des phases successives, l'organisme modifie profondément sa forme, en même temps qu'il change de mode de vie ; mais les phases permettent de réaliser ces modifications de façon progressive, en ne faisant appel qu'à des mécanismes physiologiques simples ; la croissance en plusieurs phases, chacune avec ses règles propres, offre une grande souplesse pour le passage entre les morphologie et physiologie des premiers stades et celles de l'adulte, souvent très différentes. Une série de degrés conduit des simples changements de phase aux métamorphoses impliquant des remaniements beaucoup plus profonds, souvent accompagnés de changements brusques du mode de vie. Dans les simples discontinuités, un organe se met à croître beaucoup plus vite que les autres pendant un certain temps. Dans les métamorphoses complètes, il n'y a pas seulement croissance, mais encore embryologie à retardement, avec des organogenèses complexes : un individu entièrement différent peut même se développer à partir de quelques îlots de cellules de la larve.
Le taux d'allométrie de croissance est le plus intéressant des deux paramètres de l'équation y = ax + b. Sa valeur est en effet une constante caractéristique de l'espèce et de la phase, inscrite dans le patrimoine héréditaire. L'indice d'origine est plus variable, car la place du changement de phase, qui en détermine la valeur, peut être modifiée par le milieu ; toutefois, ces variations ne prennent guère d'importance que lors de changements brusques du taux d'allométrie.
Les courbes de croissance sont donc le plus souvent utilisables, au même titre que les taux et les gradients, pour définir, rapprocher ou distinguer les espèces. Ces courbes fournissent des renseignements plus précis que les simples rapports Y/X puisque les proportions varient au cours de la croissance quand il y a allométrie. Des méthodes statistiques spéciales (analyse de variance) permettent de comparer les courbes d'allométrie, en fonction de leur écart et de la dispersion autour des segments d'allométrie des points représentatifs des individus mesurés. D'autre part, les lois de l'allométrie sont souvent suivies, chez les arthropodes en particulier, avec une précision inhabituelle en biologie, précision telle que la connaissance d'un fragment d'appendice permet de reconstituer l'animal entier, sans qu'il soit besoin de savoir même son âge.
Les méthodes biométriques et graphiques d'étude de la croissance relative ont aussi un intérêt explicatif. Elles permettent de comprendre et d'expliquer les phénomènes mis en jeu dans la croissance, et fournissent un guide à l'expérimentation dans ce domaine, même lorsque l'allométrie est suivie avec une précision médiocre ou qu'une autre loi moins simple doit être retenue. Les fœtus des singes anthropoïdes ressemblent beaucoup à celui de l'homme, et à l'homme adulte, par le faible développement de la face par rapport au crâne ; on avait cru pouvoir en déduire que l'homme dérivait du singe par fœtalisation, mais la biométrie a permis de constater que la réduction relative de la face chez l'homme s'expliquait simplement par un taux d'allométrie un peu moins élevé que chez les singes. L'allométrie a permis d'expliquer comment se produisait la féminisation des Macropodia et Inachus sous l'influence d'un parasite, la sacculine, chez ces petites araignées de mer ; la mue de puberté se marque par des discontinuités, avec une croissance des chélipèdes double de la normale chez les mâles et un élargissement brutal de l'abdomen chez les femelles ; l'étude des graphiques montre, sans qu'il soit besoin de procéder à aucune expérience, que le parasite empêche la mue de puberté chez les mâles, alors qu'il accélère son apparition chez les femelles.
Limites de la méthode
La loi d'allométrie est suivie avec une précision plus ou moins bonne suivant les animaux. Chez les arthropodes, la dispersion autour des segments d'allométrie est faible, les coefficients de corrélation sont voisins de l'unité parce que les mesures portent presque uniquement sur l' exosquelette. Chez les vertébrés, la précision reste bonne quand les os sont comparés entre eux, mais elle diminue quand on compare le squelette (dont dépend la taille) à d'autres organes. Ceux-ci peuvent réagir en effet à des facteurs qui restent sans influence notable sur le squelette. Il n'est pas étonnant en particulier que les parties du corps, qui peuvent croître et décroître une fois la taille finale atteinte, ne soient pas liées par une relation précise à la taille au cours du développement. L'hypertrophie fonctionnelle ou l'atrophie pour non-usage sont une cause de variation : un membre plâtré s'amaigrit ; un sportif acquiert par l'entraînement des biceps ou des mollets volumineux, et son cœur grossit. Les variations peuvent aussi être dues à une compétition entre divers organes.
Chez les Mollusques, les mesures se limitent souvent à la coquille pour des raisons pratiques. La croissance de cette coquille et sa forme sont souvent très sensibles aux conditions du milieu, si bien que les taux d'allométrie ne sont plus des constantes caractéristiques de l'espèce, et que la comparaison entre espèces ne peut se faire que dans un milieu donné.
Chez les animaux coloniaux ou les végétaux, l'allométrie est de peu de secours pour l'étude de la croissance. La forme de l'organisme se prête peu aux mesures et dépend surtout des endroits où peuvent apparaître les nouvelles unités (feuilles d'un arbre ou polypes d'un Cnidaire), et des corrélations qui permettent à telle unité de se développer, plutôt qu'à telle autre.
Accédez à l'intégralité de nos articles
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- André MAYRAT : sous-directeur du laboratoire de zoologie de l'École normale supérieure, Paris
- Raphaël RAPPAPORT : professeur de biologie du développement et de la reproduction, chef de l'unité d'endocrinologie pédiatrique et du diabète à l'hôpital des Enfants malades, directeur de l'unité de recherche I.N.S.E.R.M. sur la biologie de la croissance
- Paul ROLLIN : professeur à la faculté des sciences de Rouen
- Encyclopædia Universalis : services rédactionnels de l'Encyclopædia Universalis
Classification
Médias
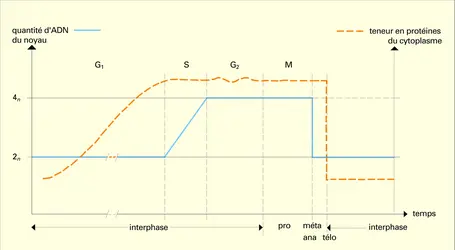
Caractéristiques de la croissance au cours du cycle cellulaire
Encyclopædia Universalis France
Rat
Encyclopædia Universalis France

Papillon
Planeta Actimedia S.A.© Encyclopædia Universalis France pour la version française.
Autres références
-
MITOSE
- Écrit par Nina FAVARD
- 6 522 mots
- 5 médias
 Pendant les périodes decroissance cellulaire (surtout la phase G1 du cycle cellulaire), le nombre des organites s'accroît en même temps que la taille de la cellule. Leurs mouvements, guidés par les microtubules, les distribuent topographiquement dans le cytoplasme dont le volume augmente. Cette distribution...
Pendant les périodes decroissance cellulaire (surtout la phase G1 du cycle cellulaire), le nombre des organites s'accroît en même temps que la taille de la cellule. Leurs mouvements, guidés par les microtubules, les distribuent topographiquement dans le cytoplasme dont le volume augmente. Cette distribution... -
AGROMÉTÉOROLOGIE
- Écrit par Emmanuel CHOISNEL et Emmanuel CLOPPET
- 6 628 mots
- 7 médias
 ...restriction hydrique sévère au cours de cette période critique peut occasionner des baisses de rendements allant jusqu'à 50 p. 100 du poids des grains. On définit également des périodes sensibles à l'égard de la croissance. Ainsi, la betterave sucrière est une plante réputée très sensible à l'eau. Toute...
...restriction hydrique sévère au cours de cette période critique peut occasionner des baisses de rendements allant jusqu'à 50 p. 100 du poids des grains. On définit également des périodes sensibles à l'égard de la croissance. Ainsi, la betterave sucrière est une plante réputée très sensible à l'eau. Toute... -
AMINOACIDES INDISPENSABLES
- Écrit par Geneviève DI COSTANZO
- 601 mots
Au début du xxe siècle, les recherches de Wilcock et de Hopkins montrèrent que certaines protéines (gélatine, zéine) sont incapables, malgré un apport azoté quantitativement suffisant, de maintenir l'équilibre nutritif de l'animal et d'assurer sa croissance. L'adjonction à ce régime de certains...
-
ANIMAUX MODES DE REPRODUCTION DES
- Écrit par Catherine ZILLER
- 4 448 mots
- 4 médias
 La reproduction asexuée ne se déclenche que lorsque l'animal atteint une certaine taille. Elle est la conséquence de la croissance. Il en est ainsi pour l'hydre, qui commence à produire des bourgeons quand la colonne gastrique, qui est en perpétuelle croissance, a une longueur suffisante.
La reproduction asexuée ne se déclenche que lorsque l'animal atteint une certaine taille. Elle est la conséquence de la croissance. Il en est ainsi pour l'hydre, qui commence à produire des bourgeons quand la colonne gastrique, qui est en perpétuelle croissance, a une longueur suffisante. - Afficher les 57 références
Voir aussi
- MALNUTRITION
- ALLOMÉTRIE, biologie
- IMMORTALITÉ POTENTIELLE, biologie
- TESTOSTÉRONE
- THYROÏDIENNES HORMONES
- MORPHOGENÈSE ANIMALE
- SACCULINE
- ENDOCRINE SÉCRÉTION
- RELEASING FACTORS (RF)
- ANTÉHYPOPHYSE
- POÏKILOTHERMIE ou PŒCILOTHERMIE
- SOMATOTROPE HORMONE (STH) ou HORMONE DE CROISSANCE ou GH (growth hormone)
- GIGANTISME
- ACROMÉGALIE
- PARATHORMONE
- CORTISOL ou HYDROCORTISONE
- GLUCOCORTICOÏDES
- HORMONOTHÉRAPIE
- ESTRADIOL ou ŒSTRADIOL
- CHITINE
- CRABE
- PUBERTÉ
- PÉRIOSTE
- VÉGÉTALE BIOLOGIE
- CORTICOÏDES ou CORTICOSTÉROÏDES
- ORGANOGENÈSE
- ENFANT MALADIES DE L'
- GRADIENT, biologie
- PÉDIATRIE
- PHYSIOLOGIE
- RENOUVELLEMENT BIOLOGIQUE
- SEXUEL COMPORTEMENT
- MÉTABOLISME CELLULAIRE
- CUSHING MALADIE DE
- SEXUELLES HORMONES
- CHROMOPROTÉINES
- MÉRÉSIS, biologie végétale
- TEMPÉRATURE
- VRILLE, botanique
- TRIIODOTHYRONINE (T3)
- HYPOTHYROÏDIE
- INSUFFISANCE ANTÉHYPOPHYSAIRE ou HYPOPITUITARISME
- DORMINE ou ABSCISSINE II
- REPRODUCTION SEXUÉE
- REPRODUCTION ASEXUÉE
- GERMEN, biologie
- CIRCUMNUTATION
- MUE
- VITAMINES D ou CALCIFÉROLS
- MÉTAZOAIRES
- ARAIGNÉE DE MER
- EXUVIATION
- CARTILAGE DE CONJUGAISON ou CARTILAGE DE CROISSANCE
- SOMA, biologie
- DIVISION CELLULAIRE
- INSULINE
- CULTURES BIOLOGIQUES
- ISOMÉTRIE, biologie
- CAMBIUMS ou ASSISES GÉNÉRATRICES ou MÉRISTÈMES SECONDAIRES
- CYTOKININES
- INDOLE-3-ACÉTIQUE ACIDE (AIA)
- QUIESCENCE
- PHLOÈME ou LIBER, botanique
- LARVE
- OSSIFICATION ou OSTÉOGENÈSE
- RÉCEPTEURS HORMONAUX
- UNICELLULAIRES
- NEUROENDOCRINOLOGIE
- CHRONOBIOLOGIE
- BERTALANFFY LUDWIG VON (1901-1972)
- PHYSIOLOGIE VÉGÉTALE ou PHYTOBIOLOGIE
- DIFFÉRENCIATION CELLULAIRE ou CYTODIFFÉRENCIATION
- EMBRYOGENÈSE ANIMALE
- DÉVELOPPEMENT HUMAIN
- MESSAGER, biochimie
- MORPHOLOGIE, biologie
- NUTRITION HUMAINE
- RÉGULATIONS BIOCHIMIQUES
- AUXÉSIS, biologie végétale
- CORTICOTHÉRAPIE
- EXOSQUELETTE
- CROISSANCE HUMAINE
- SQUELETTE HUMAIN
- FACTEURS DE CROISSANCE
- TAILLE CORPORELLE
- EXUVIE
- SOMATOMÉDINE
- IGF (insulin-like growth factor)
- ÉNANTIOMÉTRIE
- RETARD DE CROISSANCE
- POIDS CORPOREL
