- 1. Naissance de l'idée d'évolution
- 2. Darwin et la sélection naturelle
- 3. Impact et difficultés de la théorie darwinienne
- 4. Hérédité des caractères biologiques
- 5. De la crise du transformisme à la théorie synthétique
- 6. Évolution de la synthèse : remises en cause et élargissements (1970-1990)
- 7. Situation contemporaine : la « supersynthèse évo-dévo »
- 8. De l'antidarwinisme aux néo-créationnismes contemporains et à leurs conséquences
- 9. Bibliographie
ÉVOLUTION
Article modifié le
De la crise du transformisme à la théorie synthétique

Un mutant de drosophile à quatre ailes:
David Scharf/ SPL France

Au début du xxe siècle, plusieurs théories matérialistes (néo-darwinisme, néo-lamarckisme, orthogenèse, qui conçoit l'évolution des lignées « en ligne droite », et mutationnisme) sont en concurrence pour expliquer les mécanismes de l'évolution, mais chacune semble porter des coups fatals aux autres. Seule l'émergence de nouveaux modes de pensée et de nouvelles pratiques, qui aboutira à la théorie synthétique, permettra de sortir de cette crise du transformisme.
D'abord, le principe de sélection naturelle va faire l'objet d'un premier traitement opératoire, selon une approche désormais quantitative. La biométrie, développée par Francis Galton (1822-1911), va permettre de définir les paramètres statistiques nécessaires à une analyse quantitative de la sélection naturelle à l'échelle des populations.
Ensuite, depuis la redécouverte des lois de Mendel (1900), la génétique naissante apporte enfin une théorie de l'hérédité quantitative et formalisée. Celle-ci va permettre d'élaborer une théorie quantifiée et prédictive de l'évolution des gènes dans les populations, exprimée dès 1908 par la loi d'équilibre de G. H. Hardy et W. Weinberg. Selon celle-ci, une population mendélienne garde la même distribution de fréquence de ses gènes s'il n'y a ni mutation, ni migration, ni sélection naturelle.
À partir de ces données, et parallèlement à la théorie chromosomique de l'hérédité magnifiquement développée sur la drosophile par l'école de Thomas H. Morgan, s'élabore la génétique théorique des populations. Ses modèles les plus fondamentaux ont été construits au cours des années 1920 et 1930 par R. Fisher et J. B. S. Haldane en Angleterre et S. Wright aux États-Unis, ainsi que par Sergeï Tchetverikov (1880-1959) en Russie. Ces chercheurs formalisent des modèles d'évolution en examinant comment les divers facteurs (mutation, sélection, effectifs...) interagissent et peuvent modifier les populations. Ils démontrent formellement ainsi que la sélection constitue bien une « force concrète », autrement dit un mécanisme naturel efficient pouvant imposer des transformations aux populations, mais qu'elle n'est efficace que dans certaines conditions. Avec l'invention en France, par Georges Teissier et Philippe l'Héritier (1934), des « cages à populations » de drosophiles, les prévisions de la génétique des populations peuvent être testées : pour la première fois, l'évolution entre dans le domaine expérimental. La génétique des populations a été ainsi le « noyau dur » de la « synthèse moderne » ou « théorie synthétique de l'évolution » (Julian Sorell Huxley, 1942), vaste mouvement de réorganisation de toutes les disciplines scientifiques impliquées dans l'étude de l'évolution et qui se cristallise au congrès de Princeton (1947). Son ancrage disciplinaire, initialement réalisé par Theodosius Dobzhansky autour de la génétique des populations, a été complété par la systématique (Ernst Mayr, Bernhard Rensch) et par la paléontologie (George Gaylord Simpson), ces divers domaines interagissant autour de la notion d'espèce et des mécanismes de la spéciation, qui leur ont servi de « dénominateur commun » dans une perspective populationnelle.
La synthèse postule que tous les processus évolutifs doivent être compatibles avec les données de la génétique et, par ailleurs, que la sélection naturelle constitue le facteur principal orientant le changement évolutif. Sur le plan théorique, elle constitue donc fondamentalement une synthèse de la génétique et du néo-darwinisme. Pourtant, à l'origine, la théorie synthétique souhaite véritablement mériter son nom en faisant preuve, sans doute pour des raisons en partie sociologico-politiques d'« acceptabilité », d'un remarquable œcuménisme scientifique (G. G. Simpson, 1951). De la tradition darwinienne et néo-darwinienne, elle revendique la notion capitale de sélection naturelle en tant que mécanisme de la transformation évolutive par adaptation des populations. De la tradition lamarckienne et néo-lamarckienne, elle revendique l'importance de l'attention portée aux conditions du milieu et à l'adaptation corrélative des organismes et de leurs parties à celui-ci. Enfin, du mutationnisme originel, elle retient l'importance de la mutation comme cause matérielle ultime, bien qu'aléatoire, de la transformation évolutive.
Les propositions essentielles de la théorie synthétique de l'évolution dans sa forme orthodoxe (1940-1979) peuvent se résumer de la manière suivante. L'évolution observée en paléontologie, dite macro-évolution, n'est que le résultat de la micro-évolution multipliée par le facteur temps géologique. La micro-évolution (celle du généticien des populations) se réalise par des ajustements graduels, adaptatifs, des fréquences alléliques au sein des populations. En effet, dans les populations naturelles en situation « normale », il existe toujours un vaste « réservoir de variabilité » alimenté par des mécanismes génétiques (mutations géniques totalement aléatoires et recombinaisons chromosomiques) où « trie » la sélection naturelle. L'unité d'évolution étant la population, c'est au niveau de l'ensemble du génome ou « pool génique » de cette population que s'effectuent les changements significatifs imposés par la sélection. Cette dernière s'exerce finalement, au niveau des individus, comme une aptitude différentielle à la reproduction effective (notion de fitness). Dès lors, le changement évolutif est un processus lent et très graduel, et sa nature, imposée par la sélection, est adaptative. En conséquence, la macro-évolution, qui peut être reconstituée par la paléontologie, se réalise toujours au travers d'adaptations, extemporanément utiles au sein des populations, triées par la sélection naturelle et optimisées par elle.
La spéciation, c'est-à-dire l'origine des espèces, est une conséquence de l'adaptation.
L'évolution n'est pas dirigée (ce serait le finalisme), ni aléatoire (ce serait contraire à la sélection), ni quantique (ce serait peu conforme à la génétique des populations), mais graduelle, opportuniste et optimalisante.
L'ensemble de ces considérations fonde la théorie synthétique, implicitement ou explicitement, sur un certain nombre d'axiomes ou de principes généraux qu'il est possible d'expliciter comme suit :
– Principe d'utilitarisme : la sélection naturelle trie, au sein des populations, ceux des organismes individuels qui, du fait de la variabilité héréditaire spontanée, se trouvent être favorisés en termes de survie et de reproduction (fitness).
– Principe d'adaptationnisme : ce tri adapte en permanence les populations à leur environnement ;
– Principe du déterminisme sélectif : le changement évolutif étant exclusivement contrôlé par la sélection naturelle, l'adaptation est le moteur de l'évolution ;
– Principe du gradualisme (ou de répétitivité-additivité) : le changement évolutif ne peut se réaliser que très progressivement par répétition-addition de minimes modifications cumulatives ;
– Principe du réductionnisme : les variations de grande ampleur (macro-évolution) qui font sortir du cadre spécifique et engendrent les grandes nouveautés évolutives ont pour mécanismes de base (peuvent être réduites à) la variabilité intra-spécifique (micro-évolution), accumulée par la sélection naturelle ;
– Principe d'unitarisme : il n'existe ainsi qu'une seule mécanique évolutive qui, selon la durée de son action, produit la micro-évolution (intra-spécifique) dans le cadre du « temps court » (temps biologique et écologique) et la macro-évolution (trans- et supra-spécifique) dans le cadre du « temps long » (temps paléontologique et géologique). La spéciation (différenciation d'espèces nouvelles) constitue la limite entre phénomènes micro-évolutifs (intra-spécifiques), qui sont des mécanismes efficients (process), et canevas macro-évolutifs (supra-spécifiques) ou patterns, qui n'en sont que les conséquences à grande échelle.
La simplicité, la logique et la fécondité empirique de la théorie synthétique expliquent et justifient son immense succès à partir des années 1940. Une vision cohérente de l'évolution et de ses mécanismes se dégageait. Sa capacité unificatrice a résulté en partie de la stratégie « extrapolationniste » consistant à seulement voir dans l'ensemble des données macroévolutives apportées tant par la systématique que par la paléontologie des effets à grande échelle des mécanismes de base intervenant au niveau des populations. Ces derniers sont susceptibles d'approches expérimentales (cages à populations) ou, pour la spéciation, d'analyses détaillées in natura, dans toutes leurs composantes génétiques, écologiques et biogéographiques (importance des modèles d'isolement insulaire, par exemple).
Accédez à l'intégralité de nos articles
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Armand de RICQLÈS : professeur au Collège de France, chaire de biologie historique et évolutionnisme
- Stéphane SCHMITT : directeur de recherche au CNRS
Classification
Médias

Charles Darwin
Spencer Arnold/ Getty Images

Georges Cuvier
Fine Art Images/ Heritage Images/ Getty Images

Ernst Haeckel
Hulton-Deutsch Collection/ Corbis/ Getty Images
Autres références
-
ÉPIGÉNÉTIQUE ET THÉORIE DE L'ÉVOLUTION
- Écrit par Laurent LOISON et Francesca MERLIN
- 3 655 mots
- 4 médias

Le terme « épigénétique » est aujourd’hui très en vogue, aussi bien dans la littérature spécialisée que dans la presse de vulgarisation. On y associe souvent une connotation hétérodoxe et polémique, l’épigénétique étant vue comme marquant les limites de la génétique et de la...
-
EUCARYOTES (CHROMOSOME DES)
- Écrit par Denise ZICKLER
- 7 724 mots
- 9 médias
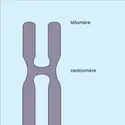 ...terminal. Tous ces individus sont interfertiles mais vivent dans des milieux différents. De telles fusions de centromères sont également invoquées dans les évolutions caryotypiques des espèces. La taille et le nombre des chromosomes ne semblent pas corrélés avec la complexité génétique des organismes : une...
...terminal. Tous ces individus sont interfertiles mais vivent dans des milieux différents. De telles fusions de centromères sont également invoquées dans les évolutions caryotypiques des espèces. La taille et le nombre des chromosomes ne semblent pas corrélés avec la complexité génétique des organismes : une... -
MITOSE
- Écrit par Nina FAVARD
- 6 522 mots
- 5 médias
 – Mitoses d' eucaryotes primitifs : l'étude des eucaryotes primitifs montre que les mécanismes de la mitose se sont compliqués au cours de l' évolution.
– Mitoses d' eucaryotes primitifs : l'étude des eucaryotes primitifs montre que les mécanismes de la mitose se sont compliqués au cours de l' évolution. -
ADAPTATION - Adaptation biologique
- Écrit par Armand de RICQLÈS
- 1 376 mots
Enfin,selon une troisième acception, l'adaptation constitue un mécanisme par lequel des transformations entre espèces peuvent intervenir au cours de l'évolution. Elle représente, là aussi, un processus dynamique. Les mécanismes mis en jeu sont typiquement ceux qui sont pris en compte par la théorie... -
AGASSIZ LOUIS (1807-1873)
- Écrit par Stéphane SCHMITT
- 1 397 mots
- 1 média

Naturaliste américain d’origine suisse, Louis Agassiz a effectué d’importants travaux en anatomie comparée et en paléontologie. Il est connu pour avoir été l’un des adversaires les plus acharnés de la théorie de l’évolution et l’un des fondateurs de la glaciologie.
- Afficher les 155 références
Voir aussi
- ÉPIGENÈSE
- FAMILLE MULTIGÉNIQUE
- OVISME
- GÈNES HOMÉOTIQUES
- THÉORIE SYNTHÉTIQUE DE L'ÉVOLUTION
- SYSTÉMATIQUE
- LYSSENKO TROFIM DENISSOVITCH (1898-1976)
- SÉQUENÇAGE, génétique moléculaire
- OWEN RICHARD (1804-1892)
- ANAGENÈSE
- CLADOGENÈSE
- EXAPTATION
- GÉNOME
- BIOLOGIE MOLÉCULAIRE
- GERMEN, biologie
- MÉTAZOAIRES
- HOMOLOGIE, phylogénie
- GRADUALISME
- HÉTÉROCHRONIE, biologie
- ZOOTYPE
- LAMARCKISME
- INFORMATION GÉNÉTIQUE
- PHYLOGÉNIE ou PHYLOGENÈSE
- CARACTÈRES ACQUIS THÉORIE DE L'HÉRÉDITÉ DES
- TRANSFORMISME
- POPULATIONS GÉNÉTIQUE DES
- ADAPTATION BIOLOGIQUE
- RÉGULATION GÉNÉTIQUE
- CRÉATION MYTHES DE LA
- EMBRYOGENÈSE ANIMALE
- GÉNÉTIQUE MOLÉCULAIRE
- CLADE
- ENSEIGNEMENT
- GERMES, biologie
- NÉODARWINISME
- CARACTÈRES, biologie
- BIOLOGIE HISTOIRE DE LA
- SCIENCES HISTOIRE DES
- BILATERIA
- RÉCAPITULATION THÉORIE DE LA ou LOI BIOGÉNÉTIQUE FONDAMENTALE
- PHYLOGÉNIE MOLÉCULAIRE
- ÉVO-DÉVO (evolutionary developmental biology)
- HOMÉOBOÎTE
- ANCÊTRE COMMUN, phylogénie
- ARBRE PHYLOGÉNÉTIQUE
- ELDREDGE NILES (1943- )
