GAMÈTES
Article modifié le
Les gamètes, aussi appelés « cellules sexuelles », sont des éléments microscopiques produits par tout individu sexué, chacun selon son sexe, et qui se rencontrent par la fécondation où ils concourent à produire ensemble un nouvel être.
Un gamète est le fruit d'une évolution cellulaire caractérisée par le déplacement, la division, et la spécialisation d'une cellule initiale appelée gonocyte. Dans l'espèce humaine les gonocytes apparaissent en surface du jeune embryon puis, vers quatre semaines, migrent vers les ébauches de glande génitale (gonade) qu'ils vont coloniser.
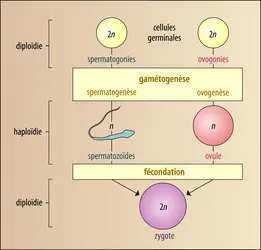
Rôle de la gamétogenèse
Encyclopædia Universalis France
À l'intérieur de la gonade, la gamétogenèse va transformer les gonocytes en gamètes. Cette évolution est complexe et comprend une forme de division cellulaire originale, la méiose, spécifique des cellules germinales. Par la méiose, le gonocyte, qui était diploïde, c’est-à-dire contenait deux lots de chromosomes, hérités l'un du père, l'autre de la mère, transmettra à chacune de ses deux cellules filles, un nombre haploïde de chromosomes, dont l’origine paternelle ou maternelle est aléatoire. Ainsi le gamète dispose d'un génome absolument nouveau et imprévisible, d'autant que des échanges entre chromosomes ou des mutations sont venus modifier les chromosomes d'origine.
Les gamètes humains
Spermatozoïdes
Les gonocytes, ayant peuplé la gonade mâle (testicule), se multiplient et donnent naissance aux spermatogonies. La transformation des spermatogonies en spermatozoïdes constitue la spermatogenèse.
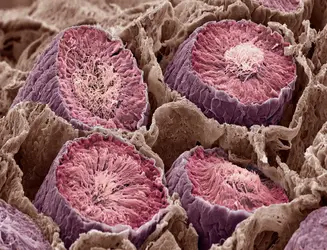
Tubes séminifères
Steve Gschmeissner/ SPL/ Getty Images
Celle-ci se déroule dans la paroi des centaines de tubes séminifères, qui occupent la plus grande partie du testicule. Entre les tubes séminifères se trouve le tissu glandulaire du testicule, contenant les cellules de Leydig qui sécrètent les hormones testiculaires, principalement la testostérone, nécessaire au déroulement de la spermatogenèse. Au sein des tubes séminifères, les cellules germinales, désormais dites spermatogonies, sont entourées par les expansions de cellules nourricières complexes, les cellules de Sertoli, lesquelles sécrètent de nombreuses protéines (inhibine, activine, Androgen Binding Protein...).

Spermatogenèse et ovogenèse
Encyclopædia Universalis France
La spermatogenèse commence à la puberté et concerne des cellules issues de la multiplication des spermatogonies : ce sont les spermatocytes de premier ordre ou spermatocytes I.
Au cours de la première division méiotique, (fig. 1) un spermatocyte I contenant quarante-six chromosomes, nombre dit 2 N parce que chaque chromosome est en double exemplaire (une paire est formée de deux homologues), dont un X et un Y, comme dans toutes les cellules du corps masculin, donne naissance à deux spermatocytes II contenant chacun vingt-trois chromosomes différents constituant l'équipement de base désigné par N (dont X ou Y selon la cellule fille). Une seconde division commence immédiatement, dans laquelle chaque chromosome se divise en deux chromatides, chacune d'elle étant destinée à l'une des deux cellules filles ou spermatides. Ainsi, théoriquement, un spermatocyte I diploïde (2 N chromosomes) est à l'origine de quatre spermatides haploïdes (N chromosomes) dont un X ou un Y selon les spermatides ; en réalité c'est seulement trois spermatides qui sont fabriquées car 25 p. 100 des cellules dégénèrent pendant cette intense activité de synthèse et réorganisation de l'ADN que renferment les chromosomes.

Spermatogenèse et spermiogenèse
Encyclopædia Universalis France
À partir de chaque spermatide va se différencier un spermatozoïde, grâce à des transformations spectaculaires qui constituent la spermiogenèse (fig. 2) et dont la finalité est de faire de cette cellule un transporteur du noyau mâle vers l'ovule qu'il féconde. Ces transformations correspondent à trois exigences complémentaires. D'abord, et afin de se déplacer aisément, le spermatozoïde va éliminer une grande partie du cytoplasme de la spermatide et condenser la chromatine contenue dans son noyau. Cette condensation est permise par le remaniement des protéines nucléaires, et particulièrement la production de protéines spécifiques au spermatozoïde, les protamines, qui se substituent aux histones chromosomiques en assurant un « empaquetage » plus serré et stable de la chromatine nucléaire. Une autre transformation essentielle est l'acquisition de la mobilité par la formation d'un f lagelle à partir du centriole distal ; ce flagelle est flanqué d'un manchon mitochondrial qui produira l'énergie nécessaire au déplacement du gamète. Enfin, l'appareil de Golgi élabore une grande vacuole riche d'enzymes protéolytiques recouvrant le noyau du spermatozoïde sous forme du capuchon acrosomique ou acrosome. Ce sac libérera son contenu enzymatique au contact des membranes de l'ovule pour permettre la pénétration du spermatozoïde.
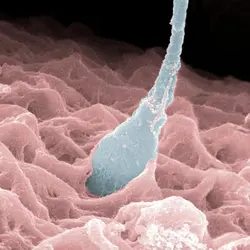
Juste avant la fécondation
clouds hill imaging ltd/ SPL/ Getty Images
L'ensemble de l'évolution du spermatocyte I au spermatozoïde se déroule dans la paroi du tube séminifère et dure soixante-quatorze jours. Puis le spermatozoïde passe dans le canal d'un organe annexe, qui coiffe le testicule, l'épididyme, où il subira des modifications plus discrètes, essentiellement de deux types, constituant sa maturation. D'une part, il acquiert son autonomie de déplacement par la mise en route du système flagellaire : le spermatozoïde, immobile lors de son arrivée dans la tête de l'épididyme, devient mobile avant d'atteindre la queue de l'organe, 6 mètres et dix jours plus tard. D'autre part, l'épithélium épididymaire est le siège de synthèses protéiques variées et de nombreuses molécules produites par l'épididyme s'attachent au spermatozoïde ; il a été montré que certaines de ces protéines interviennent dans la reconnaissance de l'ovule lors de la fécondation.

À tout moment, depuis la puberté, des spermatogonies débutent la spermatogenèse si bien que ce processus est continu, indépendant de tout phénomène cyclique. La production de spermatozoïdes, qui est maximale vers trente ans, décroît après quarante ans (sénescence), mais la spermatogenèse se poursuit cependant, grâce à la persistance de spermatogonies. Il en va tout autrement pour la femme chez laquelle la ménopause signale l'arrêt total de la gamétogenèse par épuisement du stock de cellules germinales. À l'issue de la maturation du spermatozoïde, celui-ci est d'abord stocké dans la queue de l'épididyme puis excrété dans la suite du système canaliculaire (canal déférent puis urètre), et sera émis hors du corps dans le sperme, liquide composé des sécrétions de plusieurs glandes exocrines (glandes séminales et prostate principalement).
Des facteurs de l'environnement influencent la spermatogenèse. La température du testicule est maintenue à 34 0C grâce à sa localisation dans la bourse, hors de la cavité abdominale car, à 37 0C, la spermatogenèse serait perturbée. Les radiations ionisantes (rayons X, rayons γ) lèsent les cellules germinales, surtout au niveau des spermatogonies, et ces lésions peuvent être stérilisantes. De même les antimitotiques perturbent la spermatogenèse, et la stérilité peut survenir après une chimiothérapie anticancéreuse. La pollution peut aussi altérer la fertilité masculine par diverses substances chimiques (plomb, herbicides, insecticides...) et ce phénomène est devenu très préoccupant dans la période récente puisqu'on lui attribue une diminution importante du nombre de spermatozoïdes normaux dans la semence.
En revanche, la cryoconservation du sperme (convenablement préparé à cet effet) garantit la vitalité des spermatozoïdes, permettant ainsi de les utiliser pour la fécondation in vitro du gamète femelle.
Ovules
Dans la région corticale de l'ovaire fœtal les gonocytes engendrent des ovogonies. Leur transformation en gamète féminin (ovogenèse) s'effectue au sein de massifs cellulaires, dits follicules ovariens, chaque follicule enserrant une seule cellule germinale. Contrairement à ce qui se passe pour la spermatogenèse, toutes les ovogonies, capables de se multiplier indéfiniment par mitose, ont été transformées en ovocyte de premier ordre (ovocyte I) avant même la naissance. En conséquence, le stock de cellules germinales est maximum deux mois avant la naissance – environ 5 millions – mais il n'est plus que d'un million à la naissance et ne pourra que diminuer ultérieurement. Cette diminution dépend très peu des ovulations qui surviendront dès la puberté (quelques centaines d'ovules seulement pour la vie entière), mais beaucoup plus de la dégénérescence des follicules qui se poursuit sans interruption depuis la vie fœtale. L'impossibilité de renouveler le stock des cellules germinales, contrairement à ce qui arrive chez le sujet masculin, aboutit à l'épuisement des réserves ovariennes lors de la ménopause, vers cinquante ans.
La transformation de certains ovocytes I en ovules, à partir de la puberté, est un phénomène cyclique, contrairement à la gamétogenèse masculine. Les modifications physiologiques de l'organisme pour préparer la maturation périodique d'un seul gamète sont repérables chez les femelles animales par le comportement d'œstrus qui signale la période de l'ovulation ; chez la femme, ces modifications sont repérables par les menstruations qui signalent l'absence de fécondation, deux semaines après la production d'un ovule. Ces faits montrent, autre différence avec la spermatogenèse, que l'ovogenèse s'accompagne de bouleversements hormonaux cycliques, en relation avec l'évolution des follicules. Ainsi le follicule destiné à ovuler, qui ne mesure que quelques millimètres lors des dernières menstruations, atteint 20 millimètres de diamètre en quinze jours et sécrète l'hormone œstradiol en quantités croissantes. Dès l'ovulation il se transformera en corps jaune, producteur de l'hormone progestérone, nécessaire à la grossesse. C'est le pic de sécrétion de l'œstradiol qui détermine l'œstrus chez les femelles animales, et c'est la chute du taux de la progestérone, en cas de non-fécondation, qui détermine les menstruations chez les femelles des primates comme dans l'espèce humaine.
L'ovocyte est une cellule originale de plusieurs points de vue : elle est la plus grosse cellule du corps (plus d'un dixième de millimètre de diamètre) ; elle est capable de rester au repos pendant des dizaines d'années (de la vie fœtale à la préménopause) ; elle n'est différenciée en ovule, capable d'être fécondé, que pendant quelques jours ; elle n'achève sa méiose qu'à l'issue de la fécondation ; elle contient les réserves nécessaires aux deux premiers jours du développement embryonnaire ; outre l'ADN chromosomique, équivalent à celui apporté par le spermatozoïde, elle contient dans ses mitochondries un ADN non nucléaire (mitochondrial) qui sera transmis par la mère de génération en génération. Aussi cette grosse cellule ronde, d'apparence banale, est en réalité une structure complexe dont on commence à connaître l'histoire et les propriétés.
L'ovocyte I, dérivé d'une ovogonie dans l'ovaire fœtal, est d'abord entouré de quelques cellules folliculeuses aplaties avec lesquelles il constitue un follicule primordial. Dans cet ovocyte I, la méiose a commencé mais s'est arrêtée peu après la duplication de l'ADN (au stade diacinèse de la première prophase), et le follicule restera inchangé jusqu'aux mois précédant son ovulation, ou son atrésie.
La folliculogenèse est le processus d'évolution du follicule vers sa maturité : la multiplication des cellules folliculaires produit la granulosa, qui se creuse d'une cavité liquidienne (l'antrum) et l'ensemble est entouré par deux enveloppes dont l'une, la thèque interne, contient des cellules sécrétrices d'hormones stéroïdes, surtout androgènes. Au sein d'un massif de cellules de la granulosa (le cumulus oophorus) se trouve l'ovocyte I, progressivement entouré d'une coque amorphe (la zone pellucide) composée de glycoprotéines sécrétées par l'ovocyte. C'est la gonodatropine hypophysaire F.S.H. qui induit le recrutement des follicules de la réserve et l'évolution se poursuit pendant deux mois depuis la formation de l'antrum jusqu'à l'ovulation. L'ovocyte I grandit (passant de 40 à 120 μm de diamètre) car son cytoplasme accumule des protéines (et aussi de l'ARN) qui seront utilisés dans les premiers jours du développement embryonnaire ; mais le noyau ovocytaire reste bloqué en prophase méiotique. À la maturité du follicule la granulosa compte 50 millions de cellules et l'antrum contient 5 millilitres de liquide folliculaire. C'est alors que la méiose reprend son cours sous l'influence d'une décharge, par l'hypophyse, de l'hormone lutéinisante L.H. : les jonctions unissant l'ovocyte aux cellules du cumulus se rompent, empêchant la diffusion vers l'ovocyte des facteurs folliculaires qui bloquaient sa division méiotique. Celle-ci se poursuit donc avec l'élimination, sous la zone pellucide, de la moitié des chromosomes ovulaires dans une petite masse de cytoplasme : c'est le premier globule polaire, destiné à dégénérer. En même temps se produisent des synthèses continues d'ARN et protéines qui seront nécessaires à la fécondation et au début du développement. En outre l'ovocyte synthétise à ce moment un facteur indispensable à la transformation du noyau du spermatozoïde fécondant. De nombreux grains de sécrétion issus de l'appareil de Golgi viennent se placer sous la membrane ovulaire et ces granules corticaux jouent un rôle important pour ne laisser pénétrer qu'un seul spermatozoïde lors de la fécondation. Ces événements, qui se déroulent pendant trente heures environ, constituent la maturation ovocytaire.
L'ovocyte mûr, ou ovule, présente une nouvelle phase de blocage de son matériel nucléaire, qui contient maintenant n = 23 chromosomes en métaphase de la seconde division méiotique. C'est dans cet état qu'il sera expulsé du follicule par l'ovulation et c'est la pénétration d'un spermatozoïde qui déclenchera la fin de la méiose, par l'émission du deuxième globule polaire, éliminant l'ADN résiduel qui résulte de l'achèvement du processus de réduction numérique du nombre chromosomique.
La femme ne produit normalement qu'un seul ovule par cycle menstruel mais il est possible de décupler ce nombre grâce à des traitements hormonaux, et cette pratique de superovulation est courante depuis les années 1960. L'aptitude de chaque ovule au développement est cependant réduite dans ces situations, même si la probabilité globale de procréation s'en trouve augmentée. Des anomalies du fœtus, ou de l'enfant, sont liées à des conditions mal connues de l'ovogenèse, en particulier en rapport avec l'âge des femmes, les femmes âgées concevant plus souvent des enfants porteurs d'anomalies chromosomiques telle la trisomie 21 (3 chromosomes 21 au lieu de 2).
Comme pour le gamète mâle, la cryoconservation permet de préserver les ovocytes afin de les utiliser ultérieurement (cas de personnes devant subir des traitements délétères pour l'ovaire), mais ici ce sont des fragments d'ovaire qui sont congelés. L'autogreffe, à partir d'un de ces fragments, a permis dans un cas sur deux à des patientes ainsi traitées (depuis l'an 2000) d'obtenir les naissances espérées.
Accédez à l'intégralité de nos articles
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Michel FAVRE-DUCHARTRE : professeur de botanique à la faculté des sciences, université de Reims
- Jacques TESTART : directeur de recherche honoraire à l'INSERM
Classification
Médias
Rôle de la gamétogenèse
Encyclopædia Universalis France
Tubes séminifères
Steve Gschmeissner/ SPL/ Getty Images
Spermatogenèse et spermiogenèse
Encyclopædia Universalis France
Autres références
-
MÉIOSE
- Écrit par Marguerite PICARD et Denise ZICKLER
- 4 647 mots
- 7 médias
 ...la souche sauvage présente un corps brun, et l'on connaît, entre autres, une souche mutante dont le corps est noir. Les produits de la méiose sont les gamètes, ovules ou spermatozoïdes selon le sexe, non observables directement. Cependant, le principe de ségrégation 2 : 2 d'un couple d'allèles se manifeste...
...la souche sauvage présente un corps brun, et l'on connaît, entre autres, une souche mutante dont le corps est noir. Les produits de la méiose sont les gamètes, ovules ou spermatozoïdes selon le sexe, non observables directement. Cependant, le principe de ségrégation 2 : 2 d'un couple d'allèles se manifeste... -
EUCARYOTES (CHROMOSOME DES)
- Écrit par Denise ZICKLER
- 7 724 mots
- 9 médias
 ...une alternance régulière de deux phases. Durant la phase haploïde, souvent réduite chez les organismes supérieurs aux seules cellules sexuelles ou gamètes, le noyau contient un lot de chromosomes : par exemple n = 4 chez la drosophile, n = 13 chez le Coprin, n = 10 chez le maïs, ...
...une alternance régulière de deux phases. Durant la phase haploïde, souvent réduite chez les organismes supérieurs aux seules cellules sexuelles ou gamètes, le noyau contient un lot de chromosomes : par exemple n = 4 chez la drosophile, n = 13 chez le Coprin, n = 10 chez le maïs, ... -
ANGIOSPERMES
- Écrit par Sophie NADOT et Hervé SAUQUET
- 6 134 mots
- 8 médias
 La fécondation est double chez les Angiospermes : l’un des deuxgamètes mâles issus de la division de la cellule générative fusionne avec l’oosphère pour donner un zygote, puis un embryon diploïde ; l’autre gamète fusionne avec la cellule centrale pour donner une cellule triploïde qui se divisera...
La fécondation est double chez les Angiospermes : l’un des deuxgamètes mâles issus de la division de la cellule générative fusionne avec l’oosphère pour donner un zygote, puis un embryon diploïde ; l’autre gamète fusionne avec la cellule centrale pour donner une cellule triploïde qui se divisera... -
ANIMAUX MODES DE REPRODUCTION DES
- Écrit par Catherine ZILLER
- 4 448 mots
- 4 médias
 La reproduction sexuée est caractérisée par : l'existence de deux cellules reproductrices différentes selon le sexe, le gamète mâle et le gamète femelle, qui fusionnent lors de la fécondation en produisant une cellule œuf, à l'origine du nouvel individu. Cette fusion est précédée (fig. 1), dans...
La reproduction sexuée est caractérisée par : l'existence de deux cellules reproductrices différentes selon le sexe, le gamète mâle et le gamète femelle, qui fusionnent lors de la fécondation en produisant une cellule œuf, à l'origine du nouvel individu. Cette fusion est précédée (fig. 1), dans... - Afficher les 35 références
Voir aussi
- TESTICULES
- LH (luteinizing hormone) ou HORMONE LUTÉINISANTE
- OVAIRE
- HAPLOÏDIE
- ACROSOME
- FOLLICULE OVARIEN ou FOLLICULE DE DE GRAAF
- CHLOROPHYTES
- SPERME
- ZYGOMYCÈTES
- OOSPHÈRE
- SPERMATOZOÏDE
- OVOCYTE
- RHODOPHYTES ou ALGUES ROUGES
- LIGNÉE GERMINALE
- SPERMATOGENÈSE
- SPERMATOCYTE
- OVOGENÈSE
- VÉGÉTALE BIOLOGIE
- GONOCYTE
- TUBES SÉMINIFÈRES
- ÉPIDIDYME
- REPRODUCTION SEXUÉE
- GAMÉTOGENÈSE
- SPERMATOGONIE
- OVOGONIE
- CHROMOSOMES
- OVULE
- GAMÉTOPHYTE
- OVULATION
- FLAGELLE
- ACHONTÉS
- HÉTÉROCHONTÉS
- ISOCHONTÉS
- OVULE HUMAIN
- SPERMATIDE
- SPERMIOGENÈSE
- TUBE POLLINIQUE
- MICROSPORE
