GLUCIDES
Article modifié le
Holosides
Diholosides
On désigne ainsi des glucides dérivant, théoriquement, de l'élimination d'une molécule d'eau entre deux oses sous forme cyclique. En nous limitant au cas où ces deux oses sont des aldohexoses, on prévoit un très grand nombre d'isomères, cent dans le cas le plus général. Toutefois, tous les diholosides naturels et la très grande majorité des diholosides synthétiques utilisent l'un au moins des hydroxyles hémiacétaliques (1) dans cette déshydratation.
La représentation des diholosides utilise le symbole <> ; de part et d'autre figurent les noms des oses générateurs, et, à l'intérieur, les numéros des hydroxyles utilisés dans l'anhydrisation. Le corps représenté par la formule est le D-galactopyrannose-β<14>D-glucopyrannose-α.

Il existe deux types fondamentaux de diholosides : les diholosides <1-1> appelés aussi osido-osides (ou diholosides non réducteurs) et les diholosides <1-n> ou <n-1> (n = 2, 3, 4 ou 6) appelés aussi osido-oses ou diholosides réducteurs.
En effet, les premiers ne renferment que des fonctions alcool et une fonction double acétal, ne s'ouvrant pas en milieu alcalin. Ils ne réduisent pas le nitrate d'argent ammoniacal et la liqueur de Fehling, alors que les seconds possèdent une fonction hémiacétal libre, donc réductrice.
La structure des diholosides s'établit ainsi : l'hydrolyse fournit deux oses ; dans le cas des diholosides non-réducteurs, une seule ambiguïté subsiste en ce qui concerne la structure des carbones intéressés par la déshydratation : αα′, αβ′, βα′ ou ββ′. L'additivité approchée des pouvoirs rotatoires permet de lever cette ambiguïté.
En ce qui concerne les diholosides réducteurs, on méthyle tous les hydroxyles libres, puis on hydrolyse le dérivé octométhylé. L'ose engagé en < 1 est libéré sous forme de tétra-méthyl-ose ; celui engagé en n > est libéré sous forme de tri-méthyl-ose ; l'étude de ce dernier fixe facilement la valeur de n.
Tous les diholosides sont hydrolysés par ébullition avec les acides dilués ; mais, dès la température ordinaire, l'hydrolyse peut se faire sous l'influence d'enzymes spécifiques ; c'est ainsi, par exemple, que les α-glucosidases hydrolysent tous les diholosides D-glucopyrannose-α < 1-n >... et que les β-galactosidases hydrolysent tous les diholosides D-galactopyrannose-β < 1-n >...
Si l'on engage, sous forme de diholoside, un cétohexose, le numérotage est un peu différent ; dans les cétoses, le carbone hémiacétalique est numéroté 2 ; c'est ainsi que le plus connu des diholosides (le saccharose ou sucre de canne ou de betterave) est le D-glucopyrannose-α < 1-2 > D-fructofurannose-β.
Cet holoside est non réducteur, mais le devient, après hydrolyse acide, car il est transformé en D-glucose et D-fructose, tous deux réducteurs. Cette hydrolyse fait passer le pouvoir rotatoire de + 660 à − 200 ; en effet, le D-glucose à l'équilibre (+ 520) est bien moins dextrogyre que le D-fructose à l'équilibre (− 720) n'est lévogyre. C'est la raison qui a fait donner le nom d'inversion à l'hydrolyse de saccharose. Ce terme est parfois conservé dans les cas où le pouvoir rotatoire change de valeur sans changer de signe.
La plupart des diholosides sont nettement sucrés. Les diholosides non-réducteurs ne sont pas résinifiés par les alcalis, alors que les diholosides réducteurs se comportent vis-à-vis d'eux comme les oses. Seuls les diholosides réducteurs (n ≠ 2) fournissent, avec la phénylhydrazine, des osazones.
Polyholosides
Les polyholosides constituent un vaste groupe de produits naturels très répandus chez les êtres vivants. Sous l'angle fonctionnel, ces substances jouent le rôle de réserves nutritives ou de matériaux structuraux et protecteurs.
Leurs facultés protectrices se manifestent dans les exosquelettes des Crustacés et des Insectes, les parois et les capsules de certaines bactéries et enfin les gommes et exsudats qui oblitèrent les blessures chez beaucoup de plantes. Comme matériau structural, ils contribuent à maintenir la rigidité des plantes et forment les constituants principaux de leurs mucilages, tandis que, chez les animaux, ils entrent dans la composition des cartilages et des liquides intra-articulaires.
Les facteurs responsables de la spécificité des groupes sanguins, certains facteurs antigéniques et les agents anticoagulants des tissus animaux sont également des polyholosides.
Du point de vue chimique, les polyholosides sont des produits de condensation d'oses et de dérivés d'oses unis par des liaisons glycosidiques. On distingue : d'une part, les polyholosides homogènes (homopolyholosides ou homoglucanes), constitués par un seul type ose, et très largement distribués dans la nature (cas de l'amidon, du glycogène et de la cellulose) ; d'autre part, les polyholosides hétérogènes (hétéropolyholosides ou hétéroglucanes), qui peuvent résulter de l'association de deux oses ou de dérivés d'oses distincts.
Leur structure moléculaire peut être soit linéaire comme dans la cellulose, soit ramifiée comme dans la gomme arabique, soit encore de type mixte comme dans l'amidon.
Polyholosides homogènes. Polymères du glucose (glucanes)
Les polymères du glucose sont la cellulose, l'amidon, le glycogène et les dextranes.
Cellulose
La cellulose pure est représentée par les fibres de coton qui sont des cellules dont la longueur varie entre 10 et 50 mm. Les autres sources de cellulose sont le bois, le lin, le chanvre et la paille. Les usages industriels de la cellulose se fondent sur sa nature fibreuse, qui dépend elle-même de la structure moléculaire.
La cellulose est le constituant principal de la paroi cellulaire des plantes et, comme telle, la substance naturelle la plus abondante dans la biosphère. Sa fonction biologique se rattache à ses propriétés : insolubilité, inertie chimique, rigidité. L'analyse chimique (méthylation, hydrolyse acide) a permis de montrer que la cellulose est composée de longues molécules dans lesquelles des résidus D-glucosyl sont reliés par des liaisons β (1 → 4), l'unité élémentaire étant le cellobiose.
La longueur de la chaîne moléculaire, exprimée par le nombre moyen d'unités glucosyl, représente le degré de polymérisation (D.P.) ; elle peut être évaluée par des méthodes physiques et chimiques, parmi lesquelles la mesure de la viscosité. Les résultats dépendent de l'origine de la cellulose, du traitement subi par ce matériau et de la méthode de mesure. Le D.P. de la cellulose naturelle est considéré comme supérieur à 3 000. La longueur de la chaîne semble varier d'une plante à une autre, bien que des différences puissent survenir du fait de ruptures au cours de l'isolement. Les nitrocelluloses sont employées comme explosifs et comme matières plastiques.
Amidon
De grandes quantités d'amidon sont mises en réserve par les organismes végétaux. Au microscope, l'amidon apparaît sous forme de grains. Les amidons constituent un aliment pour la plante ; ils entretiennent en effet la vie de la tige ou du tubercule pendant le repos hivernal et assurent le développement de l'embryon au cours de la germination. Les principales sources commerciales de l'amidon sont le maïs, la pomme de terre et le manioc. La plupart des amidons sont formés d'un mélange de deux constituants : l' amylose, dont la structure est linéaire, et l' amylopectine, dont la structure est ramifiée. La proportion de ces fractions est caractéristique des espèces. Ainsi, les amidons du blé et de la pomme de terre renferment 15 à 30 p. 100 d'amylose, tandis que les amidons de certaines variétés de maïs, de riz et de sorgho, en Chine, sont presque entièrement constitués d'amylopectine. En revanche, les amidons de certains pois et d'autres variétés de maïs renferment surtout de l'amylose.
Contrairement à la cellulose, les amidons peuvent être dispersés dans l'eau chaude. Mis en suspension dans l'eau et chauffés à une température critique, les grains d'amidon gonflent et forment un empois qui devient une pâte en refroidissant. L'amylose serait responsable de la gélification, alors que l'amylopectine contribuerait à donner à la pâte sa cohésion.
L'amidon est hydrolysé par les acides dilués et par des enzymes ( amylases). L'hydrolyse acide totale fournit du D-glucose, tandis que l'hydrolyse enzymatique fournit surtout un diholoside, le maltose. Des fragments de poids moléculaire plus élevé, les dextrines, sont obtenus par hydrolyse acide contrôlée et action de la température.
L'amylose est formée d'une longue chaîne d'unités D-glucosyl (environ 200 à 300) assemblées par des liaisons α (1 → 4). Une telle association moléculaire, dans laquelle les groupements −CH2OH sont placés du même côté, ne peut favoriser la structure linéaire de l'amylose, celle-ci a donc tendance à s'enrouler en hélice.
Dans l'amylopectine, les unités glucosyl sont assemblées de manière à réaliser une structure très ramifiée. Aux embranchements, la molécule de D-glucose s'attache aux autres unités, en position 6, par une liaison α (1 → 6), ou encore en position 1 et 4 : α (1 → 4). En plus de ces connexions principales existent, comme on l'a découvert ultérieurement, un petit nombre de liaisons α (1 → 3).
Les molécules ramifiées ou non ramifiées qui constituent les amidons se distinguent par la couleur des complexes apparaissant en présence d'iode qui colore l'amylose en bleu et l'amylopectine en violet. Elles peuvent être séparées par l'alcool amylique et par de nombreux composés organiques simples qui précipitent l'amylose sous forme de complexes microcristallins. La connaissance de la biogenèse de l'amidon a été améliorée par la synthèse au laboratoire de l'amylose et de l'amylopectine. On est parvenu à ce résultat en utilisant deux enzymes extraites de divers produits naturels, tels que la pomme de terre. Il semblerait que la polymérisation du D-glucose, qui donnera l'amidon des plantes et le glycogène des animaux (ainsi que la combinaison de D-glucose avec le D-fructose pour former du saccharose), soit effectuée par l'intermédiaire d'un ester phosphorique du D-glucose : l'α D-glucopyrannose 1-phosphate.
Glycogène
Très voisins des amylopectines, les glycogènes constituent les réserves glucidiques des tissus animaux. Ils sont aussi produits par les levures, les champignons et diverses algues. Comme les amylopectines, leurs poids moléculaires sont très élevés et leurs structures moléculaires sont ramifiées et formées d'unités de D-glucose. Cependant, les chaînes qui les composent sont plus nombreuses et plus courtes que celles des amylopectines. Par suite, leurs molécules sont plus compactes, plus arborescentes, plus solubles et leurs solutions moins visqueuses.
Dextranes
Les dextranes se trouvent dans les matières gluantes produites par la croissance de certaines bactéries, tel Leuconostoc mesenteroides, sur milieux saccharosés. Ils ont un poids moléculaire élevé et se composent d'unités D-glucosyl unies principalement par des liaisons α (1 → 6). Des dextranes, partiellement hydrolysés et ajoutés à une solution saline physiologique, sont employés comme succédanés du plasma sanguin dans les traitements et l'état de choc.
Polyholosides homogènes. Polymères d'autres sucres
Polymères d'autres aldoses
Les galactanes, les mannanes, les xylanes et les arabanes sont des polyholosides qui existent principalement dans les parois cellulaires des plantes. Ce sont les principaux constituants des hémicelluloses et des matières pectiques.
Polymères du fructose (fructanes)

Polyholosides
Encyclopædia Universalis France
Les fructanes constituent, tout comme l'amidon, la réserve glucidique de certaines plantes, particulièrement Composées et Graminées. L' inuline, très répandue dans de nombreux organes de plantes (artichaut, dahlia), se range dans ce groupe. Elle est composée de 30 à 40 résidus D-fructofuranosyl, unis par des liaisons β (1 → 2), et de 1 à 2 unités D-glucopyranosyl placées en bout de chaîne.
D'autres variétés de fructanes existent dans les feuilles d'herbages et sont appelées levanes. Leurs molécules sont des chaînes linéaires d'environ vingt résidus fructosyl unis par des liaisons β (2 → 6) et terminés par un résidu saccharose. Des levanes du même genre sont produits par des microorganismes, comme Bacillus subtilis.
Polymères d'osamines : la chitine
La chitine est un polyholoside des exosquelettes des Crustacés (25 p. 100 de chitine et 75 p. 100 de carbonate de calcium dans la carapace d'un crabe) et des Insectes. On la trouve aussi chez certains Champignons. La structure de la chitine rappelle celle de la cellulose ; elle consiste en de longues chaînes de résidus de N-acétylglucosamine avec liaisons β (1 → 4). La chitine est une substance inerte, difficile à hydrolyser.
Polyholosides hétérogènes
Hémicelluloses
Les hémicelluloses sont un vaste groupe de polyholosides, insolubles dans l'eau, associés à la cellulose dans les parois cellulaires des plantes. Leur nature et leur composition ne sont pas complètement établies. Elles renferment une grande quantité de xylanes, polyholosides formés d'unités D-xylopyranosyl fixées par des liaisons β (1 → 4), ainsi que diverses combinaisons de L-arabinose, D-glucose, D-galactose, D-mannose, acide D-glucuronique et acide D-galacturonique. La solubilité des hémicelluloses dans les solutions alcalines diluées permet de les séparer de la cellulose.
Matières pectiques
Situées dans les parois cellulaires et dans les ciments intercellulaires (lamelles moyennes) des tissus végétaux, les matières pectiques ont un pouvoir gélifiant élevé. On les extrait de l'écorce de citron, de la pomme et de la betterave. Elles sont formées d'acide poly-D-galacturonique, partiellement estérifié par le méthanol, et associé à des arabanes et à des galactanes.
Gommes végétales
Les gommes constituent un groupe de polyholosides à structure très complexe donnant des solutions à viscosité élevée. La gomme adragante est un exsudat produit par les Astragalus (ordre des Légumineuses). La gomme arabique, provenant de certaines espèces du genre Acacia, contient du L-arabinofuranose, du D-galactopyranose, du L-rhamnose (6-désoxy L-mannose) et de l'acide D-glucuronique.
Mucopolysaccharides
On comprend sous cette dénomination un vaste groupe de substances très importantes en physiologie et qui englobe : les acides chondroïtine-sulfuriques du cartilage, l' héparine, substance anticoagulante localisée dans les mastzellen ou mastocytes du tissu conjonctif des animaux, l'acide mucoïtine-sulfurique de la muqueuse gastrique et de la cornée, l' acide hyaluronique du tissu conjonctif, du liquide synovial, du cordon ombilical de divers tissus animaux et des capsules de certaines bactéries. Ces polysaccharides sont formés par la polymérisation d'une unité disaccharidique elle-même formée d'une osamine (D-glucosamine ou D-galactosamine) et d'un acide uronique (acide D-glucuronique le plus souvent). Les osamines portent des groupements sulfate ou acétamide. Les polysaccharides sont unis à des protéines par liaisons électrovalentielles.
Polyholosides bactériens
La spécificité antigénique de certaines bactéries est souvent liée à la possession d'une capsule plus ou moins épaisse et compacte, dont la nature est déterminée génétiquement. La purification des capsules a montré que ce sont des polyholosides de poids moléculaire élevé et de composition plus ou moins complexe. De nombreux antigènes capsulaires de pneumocoques contiennent, en dehors du glucose et du galactose, des pentoses et des sucres aminés.
La paroi bactérienne, rigide, perméable à l'eau, aux sels et aux métabolites, confère à la bactérie sa forme. Elle contient des antigènes spécifiques responsables de l'agglutination somatique type 0. Elle est indispensable à la fixation et à la pénétration des bactériophages dans la cellule. Elle est constituée par un muco-complexe composé de sucres, de lipides et d'acides aminés, dont la nature et les proportions relatives varient sensiblement suivant l'organisme, et en particulier selon qu'il s'agit de bactéries Gram-positives ou Gram-négatives. Les sucres – surtout rhamnose, arabinose, glucose, plus rarement galactose, mannose – forment 20 à 60 p. 100 du poids sec. On trouve aussi des sucres aminés : N-acétylgalactosamine et N-acétylglucosamine. La glucosamine entre dans la composition de l'acide muramique (éther-oxyde de l'acide lactique et de la N-acétylglucosamine), constituant important de la structure macromoléculaire de la paroi bactérienne. On a extrait du bacille diphtérique un polysaccharide constitué par deux molécules de D-galactose, une molécule de D-mannose et trois molécules de D-arabinose.
Accédez à l'intégralité de nos articles
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Jean ASSELINEAU : professeur de biochimie à l'université de Toulouse-III
- Charles PRÉVOST : professeur à la faculté des sciences de Paris, à l'École centrale des arts et manufactures de Paris et à l'École normale supérieure de Fontenay-aux-Roses
- Fraidoun SHAFIZADEH : Manager, Pioneering Research Department, Weyerhaeuser Cy, Seattle, États-Unis
- Melville Lawrence WOLFROM
:
regent's professor of chemistry , Ohio State University, États-Unis
Classification
Médias
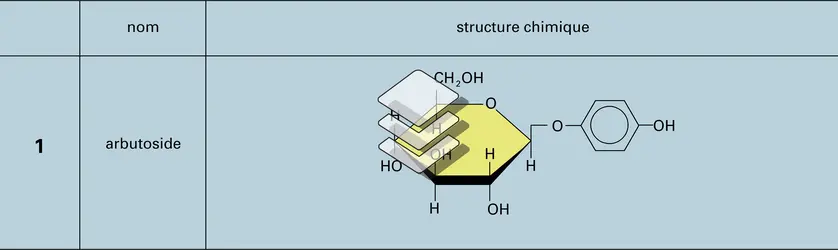
Hétérosides
Encyclopædia Universalis France
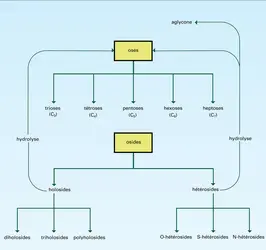
Glucides : classification
Encyclopædia Universalis France
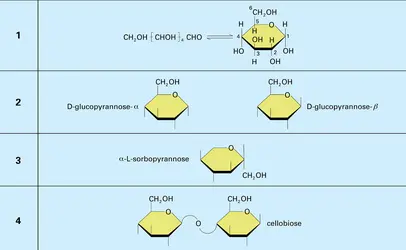
Réactions 1 à 4
Encyclopædia Universalis France
Autres références
-
ÉTUDE DES SUCRES ET DES PEPTIDES
- Écrit par Arnaud HAUDRECHY
- 208 mots
- 1 média

Élève d'Adolf von Baeyer, l'Allemand Emil Hermann Fischer (1852-1919) soutient à Strasbourg une thèse en partie consacrée à l'étude d'hydrazines, ce qui l'amène à étudier ultérieurement les sucres (glucides), à Würzburg, en 1888. Après avoir montré la structure aldéhydique du glucose,...
-
ALIMENTATION (Aliments) - Classification et typologie
- Écrit par Ambroise MARTIN
- 4 590 mots
- 1 média
Lacaractéristique nutritionnelle commune à tous ces aliments est la présence de glucides (sucres) complexes, du point de vue de leur structure chimique, et facilement digestibles, regroupés sous le terme d'amidon, organisés en granules compacts au sein des cellules végétales. La digestion... -
ALIMENTATION (Comportement et pratiques alimentaires) - Évolution de la consommation
- Écrit par Pierre COMBRIS
- 4 325 mots
- 2 médias
 ...montrent que la croissance du revenu s'accompagne d'une très forte augmentation de la part des lipides (seuls les lipides d'origine végétale régressent), d'une baisse de la part des glucides (l'accroissement de la consommation des produits sucrés ne compensant pas la baisse de la consommation des céréales)...
...montrent que la croissance du revenu s'accompagne d'une très forte augmentation de la part des lipides (seuls les lipides d'origine végétale régressent), d'une baisse de la part des glucides (l'accroissement de la consommation des produits sucrés ne compensant pas la baisse de la consommation des céréales)... -
AMINOACIDES ou ACIDES AMINÉS
- Écrit par Encyclopædia Universalis et Pierre KAMOUN
- 3 487 mots
- 6 médias
 Lescopules carbonées des aminoacides gluco-formateurs rejoignent le métabolisme glucidique et peuvent conduire au glucose puis au glycogène : cette formation de glycogène à partir des aminoacides entre dans le cadre de la néoglucogenèse qui est sous la dépendance des hormones gluco-corticostéroïdes....
Lescopules carbonées des aminoacides gluco-formateurs rejoignent le métabolisme glucidique et peuvent conduire au glucose puis au glycogène : cette formation de glycogène à partir des aminoacides entre dans le cadre de la néoglucogenèse qui est sous la dépendance des hormones gluco-corticostéroïdes.... -
ANTIGÈNES
- Écrit par Joseph ALOUF
- 7 384 mots
- 5 médias
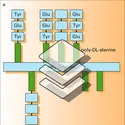 Les polyosides (polysaccharides dans la terminologie anglo-saxonne) sont des polymères à structure ordonnée, constitués par des motifs monomériques et donc des épitopes identiques se répétant à des intervalles réguliers le long de la macromolécule. Les motifs constitutifs y sont des oses ou des osides...
Les polyosides (polysaccharides dans la terminologie anglo-saxonne) sont des polymères à structure ordonnée, constitués par des motifs monomériques et donc des épitopes identiques se répétant à des intervalles réguliers le long de la macromolécule. Les motifs constitutifs y sont des oses ou des osides... - Afficher les 37 références
Voir aussi
- MOUTARDE
- GLUCOSIDES
- ENZYMES
- AMYLASES
- HYDROLYSE
- MUCOPOLYSACCHARIDES
- HYALURONIQUE ACIDE
- CATALYSE ENZYMATIQUE
- CHITINE
- NUCLÉOSIDES
- NUCLÉOTIDES
- DEGRÉ DE POLYMÉRISATION
- CAPSULE BACTÉRIENNE
- GLUCOSE-6-PHOSPHATE
- TEINTURE
- VÉGÉTALE BIOLOGIE
- ALIZARINE
- CYCLIQUES COMPOSÉS
- GLYCOGÈNE
- HÉMICELLULOSES
- CÉTONES
- ALDÉHYDES
- OXYDATION
- CELLULOSE
- MÉTABOLISME CELLULAIRE
- ALANINE
- PHOSPHORYLATION
- DÉCARBOXYLATION
- TRANSFÉRASES
- NICOTINAMIDE ADÉNINE DINUCLÉOTIDE PHOSPHATE (NADP)
- OSIDES
- XYLANE
- AMYLOPECTINE
- DEXTRANES
- CHONDROÏTINE-SULFURIQUE ACIDE
- ARBUTOSIDE
- AMYGDALINE
- INULINE
- HÉPARINE
- PECTIQUES COMPOSÉS
- SALICINE
- SAPONINES
- AGLYCONE
- MALTASE
- CELLULASES
- MÉTABOLISME DES GLUCIDES
- GLUCOSE-1-PHOSPHATE
- HEXOSES
- GLUCURONIQUE ACIDE
- OSES
- MUTAROTATION
- SORBITOL
- MANNOSE
- OSAZONES
- SORBOSE
- MALTOSE
- PYRUVIQUE ACIDE
- NICOTINAMIDE ADÉNINE DINUCLÉOTIDE (NAD)
- PENTOSES VOIE DES
- RIBULOSE
- RIBOSE
- XYLULOSE
- URIDINE DIPHOSPHATE (UDP)
- HOLOSIDES
- HÉTÉROSIDES
- ALDOSES
- CÉTOSES
- GLUCOSE
- DIASTÉRÉO-ISOMÉRIE
- GALACTOSE
- HÉMIACÉTAL
- GLYCÉRIQUE ALDÉHYDE D-
- ARABINOSE
- ÉPIMÉRISATION
- FRUCTOSE
- AMIDON
- GUANOSINE TRIPHOSPHATE (GTP)
- GLYCOGÉNOGENÈSE
- GLYCOGÉNOLYSE
- POLYSACCHARIDES ou POLYHOLOSIDES
- BIOSYNTHÈSES
- ACÉTYL-COENZYME A ou ACÉTYL-CoA
- GLUCONÉOGENÈSE ou NÉOGLUCOGENÈSE
- CYANHYDRIQUE ACIDE
- BIOÉNERGÉTIQUE
- MÉTABOLISME ÉNERGÉTIQUE
- OXALOACÉTATE
- LACTIQUE ACIDE
- CONTRACTION MUSCULAIRE
- GLYCOLYSE
- RESPIRATION
- STÉRÉO-ISOMÉRIE
- ÉNANTIOMÈRES ou INVERSES OPTIQUES ou ANTIPODES OPTIQUES
- ACÉTATES
- GLUCANES
- FRUCTANES
- LÉVANES
- DIHOLOSIDES ou DISACCHARIDES
- PHOSPHORYLASE
- CARDIOTONIQUES ou TONICARDIAQUES
- PHOSPHOÉNOLPYRUVATE (PEP)
- ALDOLASES
- URONIQUE ACIDE
- PHOSPHOFRUCTOKINASE
- MOLÉCULES BIOLOGIQUES, structure et fonction
- ALCOOLIQUE FERMENTATION
- DÉSHYDROGÉNASES
- HEXOKINASE
- PAROI CELLULAIRE
- EMBDEN GUSTAV GEORG (1874-1933)
- FIBRES VÉGÉTALES
