- 1. Anatomie et histologie
- 2. Localisations corticales sensorielles chez l'Homme
- 3. Les potentiels évoqués d'origine sensorielle et leur utilité dans la détermination des localisations corticales
- 4. Hémisphères cérébraux et motricité
- 5. Régulations motrices à partir des structures sous-corticales extrapyramidales
- 6. Hémisphères cérébraux et opérations intellectuelles
- 7. Bibliographie
HÉMISPHÈRES CÉRÉBRAUX
Article modifié le
Hémisphères cérébraux et motricité
Si l'on inclut dans les hémisphères cérébraux des Mammifères l'ensemble du télencéphale et du diencéphale, les structures qui gouvernent ou intéressent la motricité sont, d'une part, certaines aires du néocortex cérébral, d'autre part, les noyaux de la base et les régions ventrales du diencéphale qui en dépendent (substance noire et subthalamus).
Un territoire sera considéré comme moteur ou participant à l'organisation de la motricité si sa stimulation (en général électrique) entraîne des manifestations motrices (mouvements ou modifications posturales) et/ou si son ablation détermine des troubles de la motricité. La technique d'exploration électrophysiologique a permis de compléter les données de ces méthodes plus traditionnelles en précisant les mécanismes d'action des voies descendantes au niveau même des cellules nerveuses motrices (motoneurones) bulbaires ou spinales, ces dernières constituant respectivement pour la tête et le corps la voie finale commune des mouvements.

Parler de motricité sous-entend, dans la plupart des cas, les phénomènes qui relèvent de la musculature striée : en toute rigueur il s'agit de la motricité somatique, par opposition à la motricité viscérale des effecteurs végétatifs (muscles lisses, myocarde). On se limitera ici à la motricité somatique, les centres moteurs viscéraux devant faire l'objet d'une étude spéciale.
Aires motrices du néocortex des Mammifères
Aire motrice principale
On a pu délimiter, tant chez les Rongeurs que chez les Carnivores et les Primates (y compris l'Homme), une zone corticale dite aire motrice principale (ou « primaire », MI), dont les propriétés peuvent ainsi s'énoncer : sa stimulation par chocs électriques répétitifs suscite des contractions de groupes musculaires localisés de la face, des membres ou du tronc ; ces mouvements sont en règle générale controlatéraux par rapport à l'hémisphère stimulé (sauf pour la face où ils peuvent être bilatéraux) ; si la stimulation est intense, ils peuvent se prolonger au-delà du temps de stimulation (« post-décharge »).
La partie controlatérale du corps mise en jeu dépend du point cortical stimulé, ce qui a permis d'établir une somatotopie motrice (fig. 8 et 9) étonnamment semblable à la somatotopie sensitive. On y note, comme dans cette dernière, une absence de proportionnalité entre surface du corps et surface corticale correspondante ; l'importance fonctionnelle de la partie représentée (en l'espèce la finesse des mouvements exigés) dicte sans doute ces inégalités d'importance des projections, que les « figurines » tracées sur l'écorce traduisent bien par leurs distorsions.
Chez tous les Mammifères, l'aire MI est frontale, située en avant de l'aire somatique. Chez le Chat, elle occupe la majorité du cortex sigmoïde antérieur et la portion antérieure du sigmoïde postérieur, sur la face latérale et la face médiane de l'hémisphère. Chez les Primates, elle s'étend sur la circonvolution frontale ascendante, dite aussi précentrale ou prérolandique (parce que limitée postérieurement par le sillon central ou rolandique). Du point de vue cytoarchitectonique, l'essentiel de MI semble coïncider avec l'aire 4 de Brodmann, zone caractérisée par la présence de cellules pyramidales géantes (cellules de Betz) dans sa cinquième couche. Quant à sa limite antérieure, il ressort d'autres travaux qu'elle déborde assez largement l'aire 4 pour occuper la partie dorsale du champ 6 de Brodmann, où se localiserait en particulier la représentation de la musculature axiale.
De nombreuses études ont largement étendu notre connaissance du fonctionnement de l'aire MI. En abordant, sans le résoudre totalement, un vieux débat entre les tenants d'une représentation point par point en termes de muscles individuels et ceux qui concevaient l'écorce comme une « représentation de mouvements ». Les données actuelles renforcent la première conception, celle d'une mosaïque de muscles. Ainsi en est-il des recherches basées sur des stimulations de l'écorce par électrodes fines (« microstimulation », Asanuma) qui ont montré comment un certain groupe de motoneurones spinaux est commandé à partir de l'ensemble d'une colonne corticale et comment les colonnes correspondant à des motoneurones différents présentent des chevauchements partiels. D'autres analyses parlent dans le même sens lorsqu'elles décrivent les modalités complexes de décharges unitaires des neurones corticaux au cours d'un mouvement intentionnel chez le Singe (Evarts).
L'autre notion nouvelle est celle de « boucle corticale » : les explorations unitaires ont en effet révélé que les neurones de commande d'un certain groupe de motoneurones bénéficient d'afférences qui précisément proviennent du muscle commandé, ce qui confère à la motricité corticale le caractère d'une opération asservie (Phillips).
L' ablation chirurgicale partielle ou totale de MI chez le Macaque ou le Chimpanzé détermine une paralysie flasque, sans hypertonie ; cette paralysie obéit à la somatotopie puisqu'elle ne touche que les segments périphériques correspondant à la lésion. Certains réflexes, tels les réflexes abdominaux, sont déprimés ; enfin, chez le Chimpanzé et chez l'Homme, il apparaît par surcroît, après élimination de l'aire du pied, un « signe de Babinski », dorsiflexion des orteils lors de la stimulation de la sole plantaire, remplaçant la flexion ventrale normalement observée chez l'adulte éveillé. Une disparition des signes déficitaires survient après quelques semaines chez les Singes ; les séquelles sont très limitées et discrètes, les animaux redevenant en particulier capables de saisir la nourriture par opposition du pouce et de l'index. Chez le Chat, les déficits moteurs consécutifs à l'ablation de MI sont plus discrets, moins clairs à catégoriser, et la récupération plus rapide et complète encore que chez les Primates.
Aire motrice en S II
Chez les Carnivores et les Primates, un second territoire moteur distinct de MI a été identifié plus latéralement. La somatotopie y est inverse ; son extension est assez réduite. Chez le Chat, elle se localise dans les gyri suprasylvien et ectosylvien antérieur ; chez les Primates, elle occupe le pied de la circonvolution pariétale (opercule pariétal), à la limite du sillon sylvien. Dans tous les cas, cette localisation coïncide avec celle de l'aire somesthésique S II (cf. Voies corticifuges motrices, in chap. 4). Elle posséderait des projections bilatérales, en particulier pour l'aire de la face. Chez l'Homme, sa stimulation provoquerait (Penfield) tantôt une inhibition des mouvements, tantôt un « désir » de mouvement (phénomènes non obtenus à partir de MI).
Aire motrice supplémentaire
Une importante aire motrice, dite supplémentaire (AMS), a été individualisée, également chez les Carnivores et les Primates. Elle est localisée sur la face médiane où elle couvre, en avant de MI, la partie médiane de l'aire 6 de Brodmann. La représentation n'y serait pas clairement somatotopique, et les mouvements que produit sa stimulation électrique sont en général plus complexes qu'à partir de MI (mouvements d'adversion, c'est-à-dire de rotation vers le côté opposé, ou d'orientation, souvent toniques) ; ils sont parfois bilatéraux. Chez l'Homme, la stimulation de la partie de l'AMS où se projette la face ( ?) détermine en outre soit une vocalisation, soit un arrêt total de la parole. L'ablation de MI n'empêche pas l'obtention de mouvements à partir de l'AMS. Ce n'est donc pas uniquement un système de liaisons horizontales avec l'aire principale qui est essentiel à la fonction de cette aire supplémentaire, mais bien aussi une voie descendante particulière.
L'élimination de l'AMS a été effectuée systématiquement chez le Macaque. Il en est résulté des perturbations très différentes de celles qui suivent l'ablation de MI : l'animal ne présente aucune réelle paralysie, ni même de parésie (faiblesse du mouvement volontaire). En revanche, il s'installe du côté controlatéral une exagération des réflexes tendineux (tel le réflexe rotulien), une spasticité des membres (résistance exagérée à la manipulation passive) et une tendance à la préhension forcée par les doigts et les orteils (grasp). Des ablations combinées de l'AMS et de MI font apparaître un syndrome mixte : paralysie, hyperréflexie tendineuse, dépression des réflexes abdominaux, spasticité, grasp et, s'il s'agit du Chimpanzé, un signe de Babinski. Chez le Chat, les aires motrices principale et supplémentaire sont peu distantes l'une de l'autre, en sorte qu'il n'est pas aisé d'individualiser des symptomatologies différentes correspondant à la privation de chaque territoire (M. Wiesendanger, 1986).
Aires motrices oculogyres
Chez les Primates, d'autres territoires corticaux, distincts de l'AMP, suscitent des déviations conjuguées des deux globes oculaires. Cette rotation s'effectue à peu près toujours vers le côté controlatéral au cortex stimulé ; elle comporte parfois, chez l'individu éveillé, une composante verticale (c'est-à-dire que les déviations sont obliques) et, chez l'animal libre de ses mouvements, elle peut s'accompagner d'une rotation de la tête. Deux territoires oculogyres (ou plutôt oculo-céphalogyres) sont ainsi isolés. L'un est « prémoteur », coïncidant, selon des données classiques, avec l'aire 8 de Brodmann (bien que, en particulier chez l'Homme, il soit probablement plus étendu) ; l'autre est postérieur et occupe la quasi-totalité des aires occipitales (aire 17, visuelle ; aire 18, paravisuelle ; aire 19, périvisuelle). Selon Penfield, les mouvements obtenus à partir de ces aires oculo-céphalogyres ont le caractère d'activités intégrées en rapport avec l'orientation du regard (mouvements « adversifs associés »).
L'excision unilatérale du territoire oculomoteur antérieur détermine temporairement une déviation tonique des yeux et de la tête vers le côté lésé, avec difficulté de fixation dans l'hémichamp opposé. Des lésions frontales bilatérales ont entraîné un syndrome rappelant la « paralysie psychique du regard » : le regard reste fixe dans le plan médian, pour ne suivre que des points mobiles dont il se détache ensuite pour revenir à sa position initiale.
Existe-t-il d'autres territoires moteurs ?
Pour un certain nombre d'auteurs, la liste des territoires moteurs ainsi établie n'est pas close. Dès 1936, Foerster avait conclu de ses explorations neurochirurgicales chez l'Homme à l'existence de zones « motrices tertiaires » desquelles pouvaient être obtenus des mouvements adversifs grossiers. Il s'agissait en particulier de l'aire 6, dite prémotrice (en avant de l'aire 4), de l'aire 5 sur le lobe pariétal et de l'aire 22 temporale. De même E. C. Crosby (1956) et J. C. Lilly (1958) devaient élargir notablement les zones de la convexité qui, chez le Singe, peuvent être considérées comme motrices.
Ces résultats ont été très fortement mis en doute tant pour l'Homme (Penfield) que pour le Singe (Woolsey). De ces longues discussions qui n'ont pas leur place ici, dégageons cependant quelques faits essentiels.
Aire prémotrice
L'idée que le territoire situé en avant de l'aire 4, et qui occupe la partie dorsale de l'aire 6, sur la convexité représente une zone motrice particulière a été développée par tout un groupe d'auteurs. Sous le nom d'aire prémotrice, ils décrivent une région dont la stimulation déterminerait des mouvements complexes (élévations ou flexions toniques des membres ; mouvements adversifs de la tête et des membres avec participation de la musculature axiale), tandis que son ablation serait responsable de l'hyperréflexie tendineuse, de la spasticité et de la préhension forcée. D'autres travaux (Woolsey et coll., 1952, 1958), il ressort que cette région n'existe sans doute pas en tant que territoire moteur individualisé. Selon ces auteurs, en effet, l'ablation de la partie de l'aire 6 qui n'appartient ni à l'AMS ni à MI (musculature axiale) n'entraîne aucune perturbation motrice. De plus, la stimulation électrique de ce même territoire ne suscite de mouvements que pour des intensités élevées, laissant supposer qu'une diffusion du courant s'est produite soit vers l'AMS, soit vers le champ oculomoteur frontal, soit vers MI ; cela expliquerait la complexité des mouvements prétendument obtenus à partir de cette aire prémotrice et la fréquence des mouvements adversifs (S. P. Wise, 1985).
Aire somatosensitive
Certains auteurs ont estimé que chez le Singe et chez l'Homme, l'aire somatosensitive S I aurait une action motrice : sa stimulation déterminerait des mouvements selon une somatotopie semblable à la somatotopie sensitive, et ces mouvements ne seraient pas supprimés par ablation de MI, ce qui exclut le cortex moteur comme première étape dans la voie descendante. Mais la plus grande prudence s'impose actuellement à propos de ces conclusions.
Voies corticifuges motrices
Dans l'analyse du mécanisme d'action des aires corticales motrices, la connaissance des voies efférentes corticifuges s'impose. À ce titre, une distinction est à faire entre celles qui, du cortex, descendent directement dans le bulbe et la moelle, constituant le faisceau pyramidal, et d'autres, dites extrapyramidales, qui, après avoir quitté le cortex, gagnent d'abord des relais sous-corticaux pour secondairement atteindre le niveau des motoneurones.
Le tractus pyramidal
Le tractus pyramidal compte environ un million d'axones, dont 60 p. 100 sont myélinisés. En ce qui concerne son origine, il est démontré, chez les Primates et, avec des transpositions nécessaires, chez les Carnivores, que toutes les fibres pyramidales viennent du néocortex ; que le contingent essentiel est issu de l'aire 4, mais que l'aire 6, en particulier l'AMS, est à l'origine d'un contingent non négligeable ; que les cellules géantes de Betz ne constituent que 2 p. 100 des cellules d'origine du tractus ; que d'autres fibres pyramidales proviennent des aires post-centrales somatosensitives S I et probablement S II (A. M. Lassek, 1954).
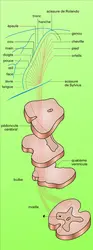
Aires motrices : voie pyramidale
Encyclopædia Universalis France
Au niveau bulbaire comme au niveau spinal, les fibres pyramidales, dont la grande majorité a traversé la ligne médiane, se terminent chez les Carnivores dans les couches dorsales et ne seront connectées aux motoneurones que par l'intermédiaire d'interneurones. Chez les Primates, un certain nombre de fibres établissent au contraire des connexions monosynaptiques avec les motoneurones. On notera de plus que des collatérales des fibres corticospinales s'achèvent dans les noyaux réticulaires bulbaires ou pontiques, et que d'autres atteignent les relais de la voie somesthésique (noyau de Goll et Burdach).
Voies corticifuges non pyramidales
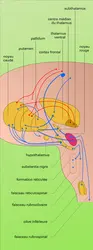
Circuits extrapyramidaux
Encyclopædia Universalis France
Divers territoires corticaux, en particulier les zones frontales, abandonnent des fibres vers des relais sous-corticaux et sont ainsi à l'origine des voies extrapyramidales : on individualise de la sorte à partir des aires 4 et 6 la voie cortico-rubro-spinale, relayant dans le noyau rouge, croisée et à distribution somatotopique comme la voie pyramidale ; la voie réticulo-spinale, avec relais dans la formation réticulée du pont et du bulbe ; la voie cortico-nigrale, vers la substantia nigra ; la voie cortico-ponto-cérébelleuse, du cortex moteur aux noyaux du pont puis à l'écorce cérébelleuse ; la voie cortico-striée, du cortex vers le striatum puis d'autres structures sous-jacentes (cf. Les structures anatomiques, in chap. 5).
On connaît mal la part respective des voies pyramidales et extrapyramidales dans les manifestations motrices observées par stimulation corticale ou les déficits résultant des lésions ou ablations. On retiendra cependant les points suivants :
– les aires 4 et 6 donnant naissance, en proportions variées, à des fibres des deux types, il n'est pas possible, en toute rigueur, d'identifier MI à une aire pyramidale, ni surtout la zone 6 à une aire extrapyramidale à distribution somatotopique ;
– chez le Macaque et le Chimpanzé toutefois, l'interruption du tractus pyramidal détermine une paralysie flasque, avec, chez le second, un signe de Babinski, mais sans aucune spasticité, ce qui évoque bien les effets de l'ablation isolée de MI ; ces observations ont été confirmées chez l'humain ;
– les répercussions d'ablations isolées de MI ou de l'AMS étant fort différentes, il ressort de l'observation ci-dessus que si MI est à dominante largement pyramidale, les voies d'action de l'AMS sont probablement plus complexes ;
– il n'est pas moins démontré qu'après section du tractus pyramidal, des mouvements peuvent encore être suscités à partir de MI, par un chemin extrapyramidal selon toute probabilité ;
– les mouvements évoqués par stimulation de l'AMS et de l'aire S I ne sont pas, on l'a vu, supprimés par ablation de MI ; cela confirme que des fibres, soit pyramidales, soit extrapyramidales et organisées somatotopiquement, transmettent cette motricité ;
– enfin, certains effets de la privation de l'AMS (spasticité, hyperréflexie tendineuse) évoquent non point une action « déficitaire », mais bien plutôt la suppression d'une influence inhibitrice exercée sans doute par un circuit extrapyramidal.
On a souvent considéré que le système pyramidal, c'est-à-dire, en schématisant, MI et le faisceau pyramidal, était indispensable à l'exécution des mouvements « volontaires ». En fait, même chez le Singe, l'élimination de ce système est suivie, après la phase parétique, d'une récupération plus ou moins complète. Si une gêne subsiste dans les mouvements fins, elle ne touche pas l'initiative motrice, mais tout au plus l'« instrument d'exécution ». Seules des ablations larges de l'ensemble du cortex moteur et prémoteur (c'est-à-dire d'une grande partie des efférences pyramidales et extrapyramidales) ont conduit à des troubles de l'initiative psychomotrice (pertes des apprentissages moteurs). Retenons aussi que chez le Macaque l'ablation subtotale bilatérale du néocortex ne supprime pas tous les signes de motricité « élaborée » (Travis et Woolsey, 1956).
Il est tout aussi probable que l' hémiplégie par lésion vasculaire prérolandique ou capsulaire chez l'Homme représente un syndrome mixte, pyramidal et extrapyramidal : car on distingue, dans son tableau clinique, d'une part la paralysie, l'abolition des réflexes abdominaux et le signe de Babinski, qui seraient d'origine pyramidale, et d'autre part la spasticité, l'hyperréflexie tendineuse qui seraient dues à une atteinte extrapyramidale.
En fait, la physiologie des aires motrices doit probablement s'interpréter en fonction d'un jeu complexe d'interactions, dont nous ignorons à peu près tout, entre les deux types de systèmes moteurs.
Accédez à l'intégralité de nos articles
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Pierre BUSER : membre de l'Académie des sciences, professeur émérite à l'université de Paris-VI-Pierre-et-Marie-Curie
- Paul LAGET : professeur de psychophysiologie à l'université de Paris-VI-Pierre-et-Marie-Curie
Classification
Médias

Champs corticaux
Encyclopædia Universalis France

Cortex cérébral : coupe en isocortex homotypique
Encyclopædia Universalis France

Cortex cérébral : cytoarchitectonie
Encyclopædia Universalis France
Autres références
-
SPÉCIALISATION ET INTÉGRATION INTERHÉMISPHÉRIQUE
- Écrit par Laura CARON-DESROCHERS et Anne GALLAGHER
- 3 032 mots
- 2 médias

C’est aux neurologues Marc Dax et Paul Broca, vers le milieu du xixe siècle, que nous attribuons les premières découvertes liées à la spécialisation hémisphérique cérébrale. En observant des patients présentant des lésions cérébrales dans l’hémisphère gauche et des difficultés langagières importantes,...
-
APHASIE
- Écrit par Jean-Luc NESPOULOUS
- 984 mots
Ainsi nommée en 1864 par le clinicien Armand Trousseau en opposition au terme aphémie retenu à la même époque par Paul Broca et au terme alalie proposé plusieurs décennies auparavant par Jacques Lordat, l’aphasie est un trouble de la production et (ou) de la compréhension du langage oral et (ou)...
-
APRAXIE
- Écrit par Didier LE GALL et François OSIURAK
- 1 288 mots
- 1 média
 ...hémisphériques gauches. Il existe cependant quelques cas dans lesquels l'apraxie idéomotrice est unilatérale, le plus souvent pour les gestes à faire du côté gauche, en particulier après une atteinte cérébrale touchant le corps calleux. D’autre part, la qualité des gestes dépend des conditions de l’examen....
...hémisphériques gauches. Il existe cependant quelques cas dans lesquels l'apraxie idéomotrice est unilatérale, le plus souvent pour les gestes à faire du côté gauche, en particulier après une atteinte cérébrale touchant le corps calleux. D’autre part, la qualité des gestes dépend des conditions de l’examen.... -
CERVEAU ET LANGAGE ORAL
- Écrit par Jean-François DÉMONET
- 2 869 mots
- 5 médias
 Les cortex temporal supérieur et frontal postéro-inférieur deshémisphères cérébraux montrent très précocement des réponses au signal de parole. Dans l’hémisphère droit, des réponses aux éléments prosodiques ont été enregistrées, alors que les régions homologues de l’hémisphère gauche montrent...
Les cortex temporal supérieur et frontal postéro-inférieur deshémisphères cérébraux montrent très précocement des réponses au signal de parole. Dans l’hémisphère droit, des réponses aux éléments prosodiques ont été enregistrées, alors que les régions homologues de l’hémisphère gauche montrent... -
CERVEAU ET PRODUCTION ÉCRITE
- Écrit par Marieke LONGCAMP et Jean-Luc VELAY
- 1 201 mots
- 1 média
 ...du cortex pariétal postérieur, autour du sillon intrapariétal. Ces observations cliniques suggèrent donc fortement que certaines parties du cerveau, dans les lobes frontal et pariétal de l'hémisphère gauche pour les droitiers, sont « spécialisées » dans l’organisation des gestes graphiques.
...du cortex pariétal postérieur, autour du sillon intrapariétal. Ces observations cliniques suggèrent donc fortement que certaines parties du cerveau, dans les lobes frontal et pariétal de l'hémisphère gauche pour les droitiers, sont « spécialisées » dans l’organisation des gestes graphiques. - Afficher les 32 références
Voir aussi
- NEUROLOGIE CLINIQUE
- HISTOLOGIE ANIMALE
- DOPAMINE
- WILLIS THOMAS (1621-1675)
- NEUROCHIMIE
- SENSIBILITÉ TACTILE ou TOUCHER
- PHYSIOLOGIE
- CORPS STRIÉS
- PRAXIES & GNOSIES
- VIEUSSENS RAYMOND (1641-1715)
- PYRAMIDALE VOIE
- PUTAMEN
- SENSORIELLES AIRES
- ROLANDO SCISSURE DE
- SYLVIUS SCISSURE DE
- AGNOSIE
- BABINSKI SIGNE DE
- MOTRICES AIRES
- HOLISTIQUE CONCEPTION
- NÉOCORTEX
- NOYAU CAUDÉ, anatomie
- NOYAU LENTICULAIRE, anatomie
- NEUROPHYSIOLOGIE
- GLOBUS PALLIDUS
- SINGES
- NEURONES DOPAMINERGIQUES
- NEURONE ou CELLULE NERVEUSE
- SUBSTANTIA NIGRA
- NIGROSTRIATAL SYSTÈME
- ANATOMO-CLINIQUE MÉTHODE
- POTENTIELS ÉVOQUÉS
- CHAT
- ANATOMIE ANIMALE
- ENCÉPHALE
- PARALYSIE
- NEUROBIOLOGIE
- STRIATUM
- NEUROCHIRURGIE
- EFFÉRENCES, neurologie
- PROJECTIONS, neurophysiologie
- AFFÉRENCES, neurologie
- STIMULUS, neurosciences
- VOIES CONDUCTRICES, neurophysiologie
- BRODMANN AIRES DE
- BETZ CELLULES DE
- SOMATOTOPIE
- DYSKINÉSIES
- MOTONEURONE ou NEURONE MOTEUR
- ANATOMIE HUMAINE
- NOYAUX GRIS CENTRAUX ou GANGLIONS DE LA BASE
- NEUROLOGIE HISTOIRE DE LA
- EXTRAPYRAMIDAL SYSTÈME
- OCULOMOTRICITÉ
- PARKINSONIEN SYNDROME
- LOBE TEMPORAL
- FRITSCH GUSTAV (1838-1927)
