
- 1. La double couche de lipides, une structure commune à toutes les membranes biologiques
- 2. Le modèle de la membrane mosaïque fluide
- 3. Mouvements affectant la membrane cellulaire
- 4. Mouvements de particules et macromolécules
- 5. Quid de la perméabilité sélective ?
- 6. La membrane comme interface dynamique
- 7. Bibliographie
MEMBRANES CELLULAIRES
Article modifié le
Le modèle de la membrane mosaïque fluide
Au moment même où le modèle de Danielli et Davson est élaboré, on sait déjà qu’il est insuffisant. En effet, par elle-même, la bicouche lipidique est essentiellement imperméable ; elle ne laisse passer facilement que les gaz, quelques substances hydrophobes, très peu l’eau et pas la majorité des molécules biologiques qui sont en règle hydrophiles. Or la cellule échange avec le milieu extérieur des centaines de molécules diverses nécessaires à la vie cellulaire. Comment le glucose traverse-t-il la membrane ? Comment la différence considérable de concentration en ions sodium (Na+) et potassium (K+) entre les deux milieux est-elle maintenue ? Comment l'influx nerveux parcourt-il l’axone et traverse-t-il les synapses ? Comment les hormones du sang peuvent-elles agir sur le métabolisme cellulaire ? Le modèle de Danielli-Davson, trop simple, ne répond pas à ces questions.
Seules des protéines insérées dans la membrane peuvent assurer des fonctions d’une telle spécificité. Danieli et Davson avaient postulé que la double couche était en sandwich entre deux couches de protéines. Ce n’est pas inexact, mais une fois encore insuffisant. Le modèle proposé par Garth L. Nicholson et Jonathan S. Singer, en 1972, résout presque parfaitement ces propositions contradictoires de perméabilité sélective et d’imperméabilité. Ce modèle conserve la double couche lipidique, mais cette dernière héberge des protéines dites membranaires. Certaines sont simplement pourvues d’une queue hydrophobe qui pénètre plus ou moins profondément la double couche. D’autres, en fait la majorité des protéines membranaires, traversent la membrane cellulaire : leur domaine extracellulaire hydrophile, presque toujours modifié par des glucides complexes, est exposé au milieu extérieur ; il est greffé sur une région hydrophobe, longue de 30 acides aminés environ, qui permet l'insertion stable de la molécule dans la membrane ; cette région hydrophobe est verrouillée sur la face interne par une courte séquence fortement hydrophile qui est, elle-même, suivie par une région hydrophile qui plonge dans le cytoplasme. Les longueurs et l'organisation des deux régions hydrophiles varient considérablement d'une protéine à une autre, d'une fonction à une autre – canaux ioniques assurant le transport contrôlé des ions, pompes à glucose, acides aminés, ions divers, pores permettant le passage de molécules sans dépense d’énergie, récepteurs des multiples messagers extracellulaires et système de transduction de signaux moléculaires ou physiques de particules (comme celles porteuses du cholestérol), récepteurs de reconnaissance cellule-cellule (comme les intégrines), etc. La membrane est ainsi peuplée d’une multitude de protéines qui assurent les différents aspects de la perméabilité sélective. On retrouve, enfin, au niveau des protéines, des éléments supplémentaires d’asymétrie membranaire : seule la fraction extracellulaire de ces protéines possède des sucres complexes ; la partie interne est en revanche glabre ou substituée transitoirement seulement par des groupements phosphate (quelques autres modifications importantes interviennent dans la transduction des signaux).
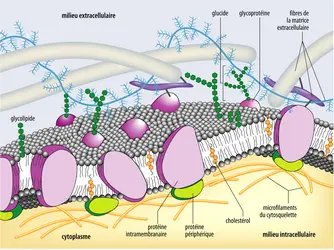
Le modèle dit de mosaïque fluide des membranes par Singer et Nicholson
Encyclopædia Universalis France
Ce modèle, dit de la membrane mosaïque fluide (parce que, vue de dessus, la membrane apparaîtrait comme des îlots de protéines flottant sur une mer de phospholipides), résout les impasses du modèle de Dianelli et Davson. Il est universellement admis et a été vérifié expérimentalement, autant par les études directes en microscopie électronique à balayage après fracturation des membranes (technique qui montre les grains de protéines dans les membranes) que par l'étude directe des protéines membranaires et par celle des fonctions qui leur sont associées.
La suite de cet article est accessible aux abonnés
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Gabriel GACHELIN : chercheur en histoire des sciences, université Paris VII-Denis-Diderot, ancien chef de service à l'Institut Pasteur
Classification
Médias

Première observation microscopique de la structure de l’écorce d’un arbre (1655)
Bettmann/ Getty Images
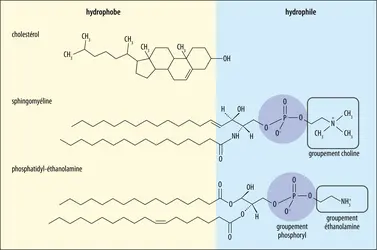
Types de lipides de la membrane cellulaire
Encyclopædia Universalis France

Visualisation de la membrane cellulaire
Biophoto Associates/ SPL
Autres références
-
MITOSE
- Écrit par Nina FAVARD
- 6 522 mots
- 5 médias
 ...Chez les cellules animales, la cytodiérèse se fait par étranglement du cytoplasme de la cellule mère (fig. 7). Au milieu ou à la fin de l'anaphase, la membrane plasmique s'invagine tout autour de la cellule selon un plan correspondant à la région équatoriale du fuseau. Le sillon circulaire ainsi formé...
...Chez les cellules animales, la cytodiérèse se fait par étranglement du cytoplasme de la cellule mère (fig. 7). Au milieu ou à la fin de l'anaphase, la membrane plasmique s'invagine tout autour de la cellule selon un plan correspondant à la région équatoriale du fuseau. Le sillon circulaire ainsi formé... -
ABSORPTION VÉGÉTALE
- Écrit par René HELLER et Jean-Pierre RONA
- 4 441 mots
- 6 médias
 Labicouche phospholipidique des membranes des cellules végétales (plasmalemme et tonoplaste) est peu perméable à l'eau et à la plupart des ions minéraux, mais leur vitesse de diffusion et la sélectivité vis-à-vis des ions peuvent être augmentées par la présence de canaux (ions, eau) et de transporteurs...
Labicouche phospholipidique des membranes des cellules végétales (plasmalemme et tonoplaste) est peu perméable à l'eau et à la plupart des ions minéraux, mais leur vitesse de diffusion et la sélectivité vis-à-vis des ions peuvent être augmentées par la présence de canaux (ions, eau) et de transporteurs... -
AQUAPORINES
- Écrit par Pierre LASZLO
- 2 344 mots
Lescellules sont encloses dans une membrane, faite d'une bicouche lipidique, qui enferme le cytoplasme, avec ses organelles et inclusions. Sur la face externe de la membrane, se trouve une autre solution aqueuse, contenant, de même d'ailleurs que le cytoplasme, des ions tels que sodium, potassium, ammonium,... -
ARCHÉOBACTÉRIES ou ARCHÉES
- Écrit par Patrick FORTERRE
- 2 138 mots
...les séquences des ARNr 16S d'archées étaient pratiquement aussi éloignées des séquences des ARNr 16S bactériens que de celles des ARNr 18S eucaryotes. L'originalité des archées a été rapidement confirmée par l'analyse de leurs glycérolipides membranaires. Alors que les glycérolipides des bactéries et... - Afficher les 49 références
Voir aussi
- INTESTIN
- RÉTICULUM ENDOPLASMIQUE
- GOLGI APPAREIL DE
- MEMBRANE PLASMIQUE ou PLASMALEMME
- PINOCYTOSE
- TRAFIC INTRACELLULAIRE
- PEPTIDE SIGNAL, cytologie
- IMMUNITAIRE SYSTÈME
- AMPHIPHILE MOLÉCULE
- PROTÉINES MEMBRANAIRES
- LIPIDES MEMBRANAIRES
- INTERNALISATION, biologie
- SODIUM, biologie
- POTASSIUM, biologie
- RÉGULATION BIOLOGIQUE
- GLYCOSPHINGOLIPIDES
- MEMBRANES BIOLOGIQUES
- PHOSPHOLIPIDES
- ORGANITES CELLULAIRES
- ANTICORPS
- MODÈLE, biologie
- PROTÉINES BIOSYNTHÈSE DES
- SPHINGOLIPIDES
- PERMÉABILITÉ, physiologie cellulaire
- CYTOLOGIE ou BIOLOGIE CELLULAIRE
- MOSAÏQUE FLUIDE MODÈLE EN
- MESSAGER, biochimie
- GLYCOLIPIDES
- GLYCOPROTÉINES
- RECONNAISSANCE, biologie
- EXOCYTOSE
- RECONNAISSANCE, immunologie
- SINGER SEYMOUR JONATHAN (1924- )
- HYDROPHOBICITÉ, chimie
- ENDOSOME
- RÉCEPTEURS MEMBRANAIRES
- LDL (low density lipoproteins)
- CYTOSQUELETTE
- CLATHRINE
- RÉCEPTEUR, biochimie
- TRANSDUCTION MEMBRANAIRE
- LIPID RAFT ou RADEAU LIPIDIQUE
- ENDOCYTOSE À RÉCEPTEURS
- ADRESSAGE DES PROTÉINES
