MOELLE ÉPINIÈRE
Article modifié le
Structure
Le seul examen à l'œil nu ou à l'aide d'une loupe à faible grossissement d'une section transversale d'une moelle épinière fraîche d'Homme ou de Mammifère permet déjà de se faire une idée assez précise des traits généraux de sa structure interne, car on y distingue deux zones : la portion centrale de couleur grise et la périphérie de couleur blanche.
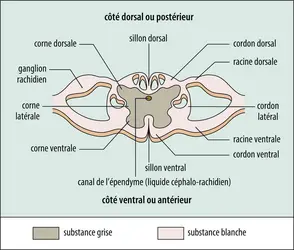
Section de la moelle épinière
Encyclopædia Universalis France
La disposition de la substance grise médullaire est très caractéristique avec ses quatre expansions symétriques qui l'ont fait comparer à un papillon. Les ailes dorsales sont dénommées cornes dorsales ou postérieures ; les ventrales, cornes ventrales ou antérieures. Les quatre cornes sont réunies par une portion centrale qui entoure un fin conduit : le canal de l'épendyme, d'un diamètre voisin du millimètre et qui demeure, dans la moelle du sujet adulte, le vestige de la lumière du tube neural embryonnaire.
Les cornes spinales dorsales sont coiffées par un étroit cordon de substance blanche, la zone ou faisceau de Lissauer, et leur configuration générale permet de distinguer en elles des portions plus ou moins renflées, dénommées : tête, col et base ; les cornes ventrales sont plus massives, surtout au niveau des renflements cervicaux et lombaires. La substance grise contient les corps cellulaires des neurones, leurs arborisations dendritiques et des fibres de longueurs et de diamètres variés, dont une partie est myélinisée. Les cornes dorsales reçoivent les fibres des racines rachidiennes correspondantes. Des cornes ventrales s'échappent les petits faisceaux de fibres qui se rassemblent à la périphérie pour constituer les racines antérieures. À tous les niveaux, des axones, dont la plupart sont myélinisés, croisent la ligne médiane entre la substance grise et le fond du sillon ventral, constituant ainsi la commissure blanche antérieure, ou ventrale.

Les cornes grises divisent la substance blanche en un certain nombre de secteurs désignés par le terme de cordon ou funiculus. On distingue ainsi : les cordons dorsaux, latéraux et ventraux. Au niveau cervical, le cordon dorsal est séparé en deux parties par le sillon dorsal intermédiaire déjà cité.
Il est possible, avec un peu d'habitude, de reconnaître le niveau d'une section médullaire transversale d'après certains traits de configuration de la substance grise. C'est ainsi que la moelle thoracique moyenne a peu de substance grise et que, à la jonction de ses cornes dorsales et ventrales, existe une expansion dénommée corne latérale. De même, le renflement cervical diffère du renflement lombaire par un développement beaucoup plus important de la substance blanche.
La moelle grise
Les neurones contenus dans la substance grise ont été tout d'abord divisés en deux groupes : les cellules radiculaires qui, par l'intermédiaire des racines antérieures, envoient leurs axones en dehors du système central, et les cellules, dites colonnaires, dont les fibres demeurent cantonnées à l'axe cérébro-spinal, certaines ne quittant pas la substance grise et formant un système d'association intrasegmentaire, les autres gagnant les différents cordons de la substance blanche.
Interprétation classique
Qu'il s'agisse des neurones de l'un ou l'autre type, on se rend compte, sur des coupes tant transversales que longitudinales, que leur répartition n'est pas homogène au sein de la moelle grise et que l'on peut y distinguer des amas, des concentrations de neurones aux contours plus ou moins nets, s'étendant sur un nombre variable de segments. On donne à ces amas le nom de noyau ou de colonne, suivant que l'on considère leur groupement tel qu'il apparaît sur des coupes transversales ou, au contraire, leur extension longitudinale. Leur nombre est élevé et leur disposition complexe ; on trouve en outre de notables différences dans leur nomenclature en fonction des auteurs et des ouvrages qui les décrivent. On ne saurait ici les citer tous et on n'indiquera que les plus importants.
Dans les cornes dorsales, la substance gélatineuse de Rolando forme une sorte de capuchon constituant la tête des cornes postérieures et s'étendant sur toute la hauteur de la moelle. À l'extrémité supérieure de celle-ci, elle se continue sans limite nette dans la formation grise du bulbe rachidien dénommée noyau spinal ou descendant du trijumeau.
Le noyau propre de la corne dorsale, qui s'étend lui aussi sur toute la hauteur de l'axe spinal, occupe, un peu plus ventralement, la région de passage entre la tête et le col.
La structure, les dimensions, la disposition des neurones et de leurs prolongements, leurs articulations synaptiques, en un mot la cyto-architectonie de ces deux structures, ont fait l'objet, depuis 1975, d'études nombreuses qui ont jeté une lumière intéressante sur leur rôle dans le contrôle et l'organisation du relais spinal des voies de la sensibilité tactile et thermo-algésique. On y reviendra plus loin.
Du huitième segment cervical au troisième segment lombaire, on trouve, dans la portion interne de la base des cornes postérieures, une colonne neuronale, facilement identifiable, comportant une proportion importante de grosses cellules : la colonne de Clarke, dont les prolongements axoniques sont le point de départ du faisceau spino-cérébelleux dorsal (direct ou de Flechsig), unissant la moelle au cervelet. La colonne de Clarke reçoit, par l'intermédiaire des racines rachidiennes postérieures, les terminaisons ou les collatérales des fibres myélinisées de gros diamètre qui véhiculent la sensibilité proprioceptive inconsciente venant des fuseaux neuromusculaires et des terminaisons nerveuses tendineuses (dites de Golgi).
Dans la portion intermédiaire qui unit les cornes dorsales et ventrales, il existe, tout à fait latéralement, entre le troisième segment thoracique et le premier segment lombaire, un amas de cellules de dimensions moyennes dont les axones sont en partie à l'origine des fibres motrices végétatives préganglionnaires du système sympathique.
Enfin, dans cette même région intermédiaire, sur la courte étendue des trois ou quatre premiers segments sacrés, le noyau parasympathique sacré (ou colonne en torsade) contient des neurones qui fournissent l'innervation parasympathique des organes pelviens : ovaires, testicules, etc.
Les cornes ventrales contiennent, entre autres, les amas de motoneurones qui assurent la commande et le contrôle de la musculature squelettique striée du tronc et des membres. Ces motoneurones sont particulièrement nombreux dans les segments médullaires qui fournissent l'innervation motrice des membres supérieurs et inférieurs, et c'est la raison de l'existence des renflements lombaires et cervico-thoraciques.
Des études très attentives, employant à la fois les techniques neuro-anatomiques et électro-neurophysiologiques les plus récentes, ont permis de repérer les noyaux motoneuronaux destinés à la commande de tel ou tel groupe musculaire et ont montré ainsi qu'ils ne se distribuaient pas au hasard mais présentaient une distribution ordonnée.
Le reste du contingent cellulaire des cornes ventrales est surtout constitué d'interneurones recevant les fibres descendantes du système extra-pyramidal (cf. infra, Principaux faisceaux descendants). Celles-ci proviennent de très nombreuses structures occupant des situations variées dans les différents étages du tronc cérébral, mais qui ont toutes en commun d'exercer un rôle très important dans l'organisation des mouvements non volontaires et le contrôle du tonus et de la posture.
Interprétation de Rexed

Substance grise médullaire : subdivision lamellaires
Encyclopædia Universalis France
La description de la moelle grise, qui se fonde sur celle des divers noyaux et colonnes, présente de nombreux inconvénients, dont la diversité des nomenclatures ; elle est en outre, par bien des aspects, assez artificielle et néglige par trop l'aspect fonctionnel des structures décrites. C'est la raison pour laquelle on préfère largement aujourd'hui utiliser la description fournie dès 1952 par B. Rexed, neuro-anatomiste suédois qui a montré que l'axe gris spinal avait une structure lamellaire, distinguant, à partir de la pointe de la corne postérieure, neuf couches ou lames successives assez bien différenciées par divers traits architectoniques. Certaines lames ou portions de lames contiennent naturellement tout ou partie des colonnes ou noyaux médullaires précités ; d'autres n'avaient pas été reconnues jusqu'alors comme des entités distinctes dans les autres modes de description.
La moelle blanche
Les divers cordons médullaires contiennent un très grand nombre de faisceaux de fibres d'importance variée et qui peuvent essentiellement être distingués par le sens des influx qui les parcourent, c'est-à-dire préférentiellement ascendants et réunissant les divers segments médullaires aux structures du tronc cérébral ou, au contraire, préférentiellement descendants, assurant la liaison de ces mêmes formations et du cortex cérébral avec les neurones spinaux. Ces faisceaux, que l'on peut caractériser comme longs, sont décrits ci-après.
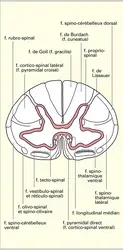
Substance blanche médullaire : faisceaux
Encyclopædia Universalis France
On doit les distinguer des voies « proprio-spinales » qui montent et qui descendent sur un nombre varié de segments médullaires, assurant d'étroites connexions entre ceux-ci et jouant un rôle essentiel dans l'organisation des réflexes plurisegmentaires ; les fibres composant ce système peuvent être classées en deux groupes : celui du faisceau dorso-latéral ou de Lissauer, déjà signalé, et celui du faisceau fondamental ou faisceau propre qui entoure d'une mince couche blanche la périphérie de la substance grise et qui est composé de fibres amyéliniques et myélinisées fines entrant ou sortant de la moelle à tous les niveaux.
Principaux faisceaux ascendants
Les faisceaux ascendants transmettent les messages venant directement de la périphérie et recueillis par certaines terminaisons sensitives cutanées ou ceux qui, d'origines diverses, également sensitives, ont au préalable relayé par différents noyaux et colonnes des cornes postérieures. Leur multiplicité apparaît souvent déconcertante pour le non-spécialiste, et leur énumération systématique est fastidieuse et finalement peu riche de signification fonctionnelle. En fait, les descriptions modernes s'appuient sur une argumentation à la fois phylogénétique, tirée de la neuro-anatomie comparée, neurophysiologique, issue des travaux récents et neuropathologique, fournie par la description des symptômes observés dans les lésions et affections médullaires. Elles reconnaissent ainsi trois systèmes essentiels, à signification fonctionnelle bien distincte :
– Les voies spino-cérébelleuses comprennent le faisceau spino-cérébelleux dorsal (direct ou de Flechsig) et le faisceau spino-cérébelleux ventral (croisé ou de Gowers). C'est par eux que les messages provenant des récepteurs de la proprioception inconsciente (fuseaux neuromusculaires et récepteurs tendineux de Golgi) gagnent le cervelet après avoir, suivant le cas, relayé dans la colonne de Clarke ou dans certaines cellules de la base des cornes ventrales.
– Les voies dites « lemniscales », d'apparition phylogénétique récente (on ne les rencontre guère que chez les Mammifères), sont constituées des faisceaux des cordons postérieurs (faisceau de Goll ou fasciculus gracilis et faisceau de Burdach ou fasciculus cuneatus) et du faisceau spino-thalamique latéral, que l'on peut dénommer aussi néo-spino-thalamique.
Les messages transportés par les faisceaux des cordons postérieurs proviennent directement de certaines terminaisons cutanées à seuil d'excitabilité bas, fortement discriminatives, voyageant dans des fibres centripètes myélinisées de fort diamètre (8 à 12 μm). Les corps cellulaires de ces dernières siègent dans les ganglions rachidiens, et les axones qui en sont issus gagnent, par la voie des cordons postérieurs, le bulbe rachidien où ils se terminent dans les noyaux de Goll et de Burdach. Les fibres qui composent les faisceaux gracilis et cuneatus sont ordonnées d'une manière remarquable : celles qui passent par les racines rachidiennes les plus basses sont les plus internes ; celles qui arrivent par les plus hautes sont les plus externes.
Le faisceau spino-thalamique latéral est fait d'axones issus en particulier de neurones du noyau propre de la corne dorsale contralatérale. Avant de le rejoindre, ils ont franchi la ligne médiane par la voie de la commissure blanche antérieure. C'est la grande voie spinale de la sensibilité thermique et de la sensibilité douloureuse, mais pour cette dernière on admet aujourd'hui qu'il ne s'agit que de son mode « rapide », c'est-à-dire de la douleur brève, bien localisable et dont la charge affective est peu intense, telle qu'on peut la ressentir par exemple à la suite d'une piqûre ou d'une coupure superficielle. Après sa traversée spinale, le faisceau spino-thalamique latéral monte dans le tronc cérébral et gagne le thalamus, où il se termine dans un complexe nucléaire dit ventro-postéro-latéral, auquel aboutissent également les fibres originaires des noyaux de Goll et de Burdach, fibres qui transportent les mêmes messages tactiles que les faisceaux spinaux portant ce nom.
– Les voies extra-lemniscales sont beaucoup moins bien systématisées, leur origine phylogénétique est ancienne, leur localisation est plus diffuse, la place tenue dans leur composition par les fibres amyéliniques et myélinisées fines est plus grande. Leur terminaison se fait dans des noyaux et formations variés du tronc cérébral et au niveau du thalamus dans des noyaux distincts du complexe ventro-postéro-latéral, précédemment cité.
On mentionnera particulièrement parmi les nombreux faisceaux (f. spino-tectal, f. spino-vestibulaire, f. spino-réticulaire...) qui composent ce système le faisceau spino-thalamique ventral (paléo-spino-thalamique) qui est situé ventralement par rapport au spino-thalamique latéral. Ses fibres, beaucoup plus nombreuses que celles du néo-spino-thalamique, sont émises à partir de neurones siégeant dans les cornes postérieures, recevant des afférences radiculaires de petit diamètre ; elles sont mises en activité par des stimuli variés : tactiles, thermiques, douloureux... Pour ces derniers, le type de sensation correspondant paraît être la douleur dite lente, profonde, pénétrante, diffuse, mal localisable, entraînant des réactions végétatives notables (pâleur, nausées, etc.) et le plus souvent, si ce n'est exclusivement, d'origine viscérale (douleur des coliques hépatiques, ou des infections péritonéales, par exemple).
Ces faisceaux, dits extra-lemniscaux, se retrouvent facilement chez les Vertébrés inférieurs, où ils constituent les seules voies médullaires ascendantes, les voies lemniscales n'étant pas encore apparues.
Principaux faisceaux descendants
Les faisceaux descendants peuvent être rangés en deux grands groupes : d'une part, le contingent d'origine corticale dont l'apparition est récente, puisqu'elle coïncide avec celle du cortex moteur cérébral qui n'existe que chez les Mammifères, et que l'on désigne du terme général de système pyramidal ; d'autre part, tous les autres faisceaux descendants rangés sous le vocable général de système extra-pyramidal.
Les faisceaux cortico-spineux ont suscité les recherches les plus nombreuses et les plus soigneuses, en raison de leur importance fondamentale dans le contrôle de la motricité volontaire chez l'Homme. Ils sont issus en particulier des grandes cellules pyramidales du cortex cérébral moteur et leurs fibres descendent dans la capsule interne homolatérale, gagnent le bulbe rachidien, s'entrecroisent sur la ligne médiane (décussation pyramidale) à la base de celui-ci, pour la plus grande part, si ce n'est la totalité, puis descendent dans la moelle en venant se ranger contre le bord antérieur des cornes postérieures et se terminent dans les cornes ventrales du même côté (faisceau cortico-spinal latéral ou faisceau pyramidal croisé). Un petit contingent d'axones, n'ayant pas subi de décussation au niveau bulbaire, va (seulement chez l'Homme et les Primates) voyager dans le cordon médullaire ventral homolatéral, le long du bord du sillon antérieur, puis se terminer dans les cornes ventrales du côté opposé, après avoir croisé dans la commissure antérieure (faisceau cortical spinal ventral ou faisceau pyramidal direct de Turck).
Les faisceaux extra-pyramidaux sont nombreux, leur systématisation est complexe. On peut les classer en fonction de leur lieu d'origine principal et distinguer ainsi ceux qui prennent naissance dans le mésencéphale de ceux qui sont originaires de structures rhombencéphaliques. Dans les premiers, on range les faisceaux ou systèmes : rubro-spinal, tecto-tegmento-spinal et longitudinal médian (bandelette longitudinale postérieure). Dans les seconds, on trouve les faisceaux ou systèmes : unciné (cérébello-spinal), réticulo-spinal, vestibulo-spinal et olivo-spinal. Certains sont principalement croisés en proportion sensiblement égale. Ils sont évidemment d'origine phylogénétique plus ancienne que les faisceaux cortico-spinaux, et on retrouve la plupart d'entre eux chez les Vertébrés inférieurs, même les plus primitifs. On se limitera ici à des indications sommaires sur leur description et leur localisation. On insistera, en revanche, sur certaines données récentes concernant les rôles respectifs et les lieux de terminaisons des deux systèmes pyramidaux et extra-pyramidaux.
Chez l'Homme et les Singes anthropomorphes, les faisceaux cortico-spinaux représentent la grande voie motrice centrale, assurant la totalité du contrôle volontaire sur les motoneurones spinaux avec lesquels d'ailleurs leurs terminaisons contractent des rapports synaptiques directs.
Quand il s'agit des autres ordres de Mammifères, on s'aperçoit que, au fur et à mesure que l'on descend vers des groupes de moins en moins perfectionnés, l'importance des faisceaux cortico-spinaux décroît peu à peu par rapport à celle des tractus extra-pyramidaux, en particulier au profit du faisceau rubro-spinal (issu d'une structure mésencéphalique, le noyau rouge, dont le rôle dans le contrôle de la motricité involontaire est essentiel) ; c'est ainsi que les faisceaux pyramidaux ont totalement disparu chez les Ongulés (Ruminants, Équidés, Porcins, Proboscidiens). D'autre part, chez les Mammifères infra-Primates – et cette constatation n'a pas manqué de prime abord de surprendre –, les terminaisons des fibres cortico-spinales ne se font plus directement sur les motoneurones des cornes antérieures, mais uniquement sur des interneurones, là ou parviennent aussi les ramifications finales des faisceaux extra-pyramidaux. Ces interneurones peuvent se situer non seulement dans les cornes ventrales, mais aussi dans la portion intermédiaire et même dans les cornes dorsales. Ces constatations anatomiques, renforcées par les observations électrophysiologiques, ont profondément modifié nos conceptions sur le contrôle cortical moteur chez les Mammifères autres que l'Homme.
Accédez à l'intégralité de nos articles
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Paul LAGET : professeur de psychophysiologie à l'université de Paris-VI-Pierre-et-Marie-Curie
Classification
Médias
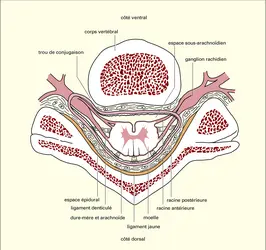
Situation au sein du canal vertébral
Encyclopædia Universalis France
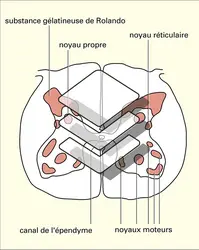
Substance grise et noyaux
Encyclopædia Universalis France
Section de la moelle épinière
Encyclopædia Universalis France
Autres références
-
BELL sir CHARLES (1774-1842)
- Écrit par Jean-Gaël BARBARA
- 1 500 mots
- 1 média
 ...nerfs jusqu’au cerveau selon leurs trajets spécifiques. Son approche est la dissection. Il se convainc que les nerfs moteurs remontent jusqu’au cerveau par les racines antérieures de la moelle épinière, et les nerfs sensitifs jusqu’au cervelet en passant par les racines postérieures de la moelle.
...nerfs jusqu’au cerveau selon leurs trajets spécifiques. Son approche est la dissection. Il se convainc que les nerfs moteurs remontent jusqu’au cerveau par les racines antérieures de la moelle épinière, et les nerfs sensitifs jusqu’au cervelet en passant par les racines postérieures de la moelle. -
BROWN-SÉQUARD CHARLES ÉDOUARD (1817-1894)
- Écrit par Encyclopædia Universalis
- 328 mots
Physiologiste et neurologue, pionnier de l'endocrinologie et de la neurophysiologie, Brown-Séquard fut parmi les premiers à étudier la physiologie de la moelle épinière. Il décrivit notamment un syndrome qui porte son nom (il correspond à l'hémisection médullaire, dont les effets...
-
CARTOGRAPHIE CELLULAIRE DU CERVEAU
- Écrit par Jean-Gaël BARBARA
- 2 990 mots
- 2 médias
...morphologiquement. C’est ainsi que l’une des premières applications de l’atlas Allen de souris a consisté à caractériser un nouveau type de cellule de la moelle épinière (des cellules de la glie radiale) se comportant comme des cellules souches et qui pourrait être ciblé pour une réparation physiologique... -
CERVEAU HUMAIN
- Écrit par André BOURGUIGNON , Cyrille KOUPERNIK , Pierre-Marie LLEDO , Bernard MAZOYER et Jean-Didier VINCENT
- 12 789 mots
- 9 médias
 ...arrière, par le mésencéphale, le pont et le bulbe rachidien. Ce dernier rejoint, en passant par le trou occipital, le canal rachidien et se prolonge par la moelle épinière. Celle-ci est parcourue par des voies descendantes, qui acheminent les signaux électriques du cerveau vers les neurones moteurs, et des...
...arrière, par le mésencéphale, le pont et le bulbe rachidien. Ce dernier rejoint, en passant par le trou occipital, le canal rachidien et se prolonge par la moelle épinière. Celle-ci est parcourue par des voies descendantes, qui acheminent les signaux électriques du cerveau vers les neurones moteurs, et des... - Afficher les 21 références
Voir aussi
- PHYSIOLOGIE
- PYRAMIDALE VOIE
- SUBSTANCE GRISE
- SUBSTANCE BLANCHE
- NEUROPHYSIOLOGIE
- NERF RACHIDIEN
- FAISCEAUX ASCENDANTS, anatomie
- FAISCEAUX DESCENDANTS, anatomie
- NEURONE ou CELLULE NERVEUSE
- BURDACH FAISCEAU DE
- GOLL FAISCEAU DE
- NEUROBIOLOGIE
- CORDONS SPINAUX
- CÉRÉBELLEUSES VOIES
- FAISCEAUX SPINO-CÉRÉBELLEUX
- MOTONEURONE ou NEURONE MOTEUR
- ANATOMIE HUMAINE
- EXTRAPYRAMIDAL SYSTÈME
