PHÉOPHYCÉES ou ALGUES BRUNES
Article modifié le
Les Phéophycées, ou Algues brunes, sont presque exclusivement marines. Par leur taille et par leur abondance, certaines d'entre elles jouent un rôle très important dans la végétation des diverses mers. Les peuplements étendus de Laminariales abritent une faune variée de poissons et de crustacés comestibles qui y trouvent une nourriture abondante. Outre les utilisations industrielles, il y a lieu de noter qu'au Japon certaines Laminaires sont largement exploitées pour l'alimentation humaine sous le nom de kombu.
Bien que leur anatomie et leur mode de reproduction soient très variés, les Phéophycées possèdent une grande homogénéité dans leur structure cytologique. Comme chez tous les Chromophytes, leurs plastes, colorés en brun par des pigments spéciaux (fucoxanthine), leur donnent une coloration brune ou olivâtre caractéristique ; mais contrairement aux autres Chromophytes, elles ne sont jamais unicellulaires et leurs cellules reproductrices mobiles (zoïdes) possèdent deux flagelles, l'un antérieur, l'autre postérieur, insérés sur le côté de la cellule.
Caractères généraux
Cytologie

Caractéristiques cytologiques
Encyclopædia Universalis France

Les cellules des Phéophycées sont généralement uninucléées, le noyau étant situé au centre de la cellule et maintenu en place par de nombreux tractus cytoplasmiques anastomosés. Les cellules possèdent un ou plusieurs plastes généralement discoïdes ou bilobés, pourvus de chlorophylle a et de chlorophylle c associées à des caroténoïdes variés dont certains leur sont propres (fucoxanthine). L'ultrastructure des plastes rappelle celle des Chromophytes ; les saccules aplatis renfermant les pigments (thylacoïdes), au lieu d'être isolés les uns des autres au sein du stroma comme chez les Rhodophycées, sont groupés généralement par deux ou trois. Les plastes de beaucoup de Phéophycées possèdent des pyrénoïdes en forme de poire faisant saillie à l'extérieur du plaste, tout en restant entourés par la membrane plastidiale : ils sont généralement de structure homogène et, contrairement à ceux des Chlorophycées, toujours dépourvus d'une enveloppe d'amidon. Les pyrénoïdes n'existent que dans les formes peu évoluées (Ectocarpales, Chordariales, Dictyosiphonales) et font défaut dans les formes plus évoluées (Dictyotales, Laminariales). Toutefois, la microscopie électronique a clairement mis en évidence un pyrénoïde dans les plastes des Sphacélariales ainsi que dans ceux des oosphères de certaines Fucales où ils étaient passés inaperçus au microscope photonique.
Les cellules de Phéophycées, toujours dépourvues d'amidon, renferment comme produit de la photosynthèse un polyholoside donnant du glucose par hydrolyse (laminarine) et un polyalcool à six atomes de carbone (mannitol). Enfin, leur cytoplasme, surtout autour du noyau, possède de petits corps réfringents incolores mais facilement teintés par les colorants vitaux, les physodes, considérés comme des vacuoles spécialisées ; ils contiennent des substances diphénoliques voisines des tannoïdes (phloroglucinol).
Les cellules reproductrices flagellées, zoogamètes ou zoospores, que l'on qualifie indistinctement de zoïdes, sont caractérisées par leurs flagelles hétérocontés, insérés latéralement et dirigés l'un vers l'avant, l'autre vers l'arrière de la cellule. Le flagelle antérieur, plus long (sauf chez les Fucales), est seul pourvu de mastigonèmes.
Morphologie
Il n'existe pas de Phéophycées unicellulaires. Les formes les plus simples se présentent sous l'aspect de filaments ramifiés formés de files de cellules toutes semblables ou souvent amincies à leur extrémité en formant des pseudo-poils (Ectocarpales). Ces filaments peuvent parfois différencier des rameaux courts latéraux (pleuridies) ou s'associer en faisceaux pour former des cladomes multiaxiaux (Chordariales).

Structures et types de croissance
Encyclopædia Universalis France
Dans les formes relativement simples (Ectocarpales), les cellules des filaments axiaux ne sont capables que de divisions transversales et la structure est dite haplostique. Ailleurs, les cellules axiales se divisent par des cloisons longitudinales, donc perpendiculaires aux cloisons transversales (Sphacélariales, etc.) ; la structure de l'axe est alors polystique .
La croissance des filaments peut être intercalaire (Ectocarpales) : certaines cellules se divisent activement par des cloisons transversales, pour donner à la fois dans les deux directions opposées de nouvelles cellules axiales semblables. Parfois, cette croissance intercalaire se localise vers le sommet des filaments axiaux : les cellules formées vers le bas constituent le filament axial et portent des pleuridies ou des axes secondaires ; tandis que, vers le haut, les cellules formées, plus minces, s'allongent en un poil terminal qui se détruit progressivement à son extrémité distale : la croissance est alors trichothallique (Chordariales). Enfin, la croissance peut être terminale, grâce à une cellule initiale qui donne naissance aux divers tissus du thalle (Dictyotales, Sphacélariales, Fucales).
Un cas particulier de croissance intercalaire est celui des Laminariales, où il existe une zone de croissance méristématique située à la jonction du stipe cylindrique et de la lame. Le fonctionnement de ce méristème stipo-frondal détermine, d'une part, la croissance et l'élargissement de la lame qui se détruit à sa partie distale soit progressivement soit à chaque reprise de la végétation au printemps (cf. algues, pl. hors texte : Laminaria hyperborea).
La croissance peut également être marginale grâce à l'activité synchrone de filaments à croissance trichothallique soudés latéralement (Cutlériales) ou de cellules initiales terminales (Padina).
Reproduction
La nature des organes reproducteurs permet de distinguer parmi les Phéophycées deux grandes sous-classes, les Phéosporées et les Cyclosporées.

Ectocarpus : cycles de reproduction
Encyclopædia Universalis France
Chez les Phéosporées, il existe deux sortes d'organes reproducteurs (zoïdocystes) produisant des cellules reproductrices flagellées : zoospores ou zoogamètes, que l'on confond généralement sous le nom de zoïdes. Les zoïdocystes uniloculaires, de forme ovoïde ou subsphérique, renferment de nombreux zoïdes entassés « comme des noix dans un sac », tandis que, dans les zoïdocystes pluriloculaires, la cellule mère du zoïdocyste se divise par des cloisonnements successifs en de nombreuses petites logettes dont chacune renferme un seul zoïde.
C'est dans les zoïdocystes uniloculaires que s'effectue la méiose ; les zoïdes produits sont donc en principe haploïdes. Il n'y a, par contre, jamais de méiose lors de la formation des zoïdocystes pluriloculaires, dont les zoïdes ont le même nombre de chromosomes (haploïde ou diploïde) que l'individu qui les produit. Ces zoïdes se comportent soit comme des spores, soit comme des gamètes. Dans ce dernier cas, il y a tantôt isogamie, les gamètes étant produits dans des zoïdocystes tous semblables, tantôt anisogamie plus ou moins prononcée, les gamètes mâles étant alors produits dans des zoïdocystes à petites logettes et les gamètes femelles dans des zoïdocystes à grandes logettes. Parfois aussi, il y a oogamie et les gamétocystes femelles ne renferment qu'un seul gamète.

Fucus vesiculorosus : organes reproducteurs
Encyclopædia Universalis France
Chez les Cyclosporées (Fucales), la reproduction est toujours oogame ; les gamètes femelles immobiles (oosphères) se forment dans des oogones et les gamètes mâles (anthérozoïdes) dans des spermatocystes. Ces deux types d'organes sont inclus dans des conceptacles creusés dans des parties différenciées de la fronde (réceptacles).
Cycles d'alternance de générations

Laminaria : prothalles
Encyclopædia Universalis France
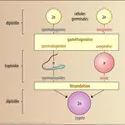
Chez les Phéosporées, le cycle est, en principe, digénétique, diplohaplophasique, avec alternance d'un gamétophyte haploïde et d'un sporophyte diploïde. Dans les formes les moins évoluées (Ectocarpales), gamétophyte et sporophyte sont morphologiquement identiques ; parfois aussi, il existe entre les deux générations un dimorphisme très net (Cutlériales), mais, dans tous les cas, le gamétophyte présente une structure au moins aussi différenciée que le sporophyte. L'ensemble de ces Algues constitue le groupe des Isogeneratae, par opposition aux Heterogeneratae (Laminariales) dont le gamétophyte, microscopique et réduit à des filaments plus ou moins ramifiés, constitue un prothalle.
Toutefois, chez beaucoup d'espèces (Ectocarpales, Chordariales et Dictyosiphonales), le cycle peut devenir très compliqué du fait que le même type d'organe reproducteur (uniloculaire ou pluriloculaire) peut se rencontrer aussi bien sur des individus haploïdes que diploïdes ; selon les cas, il peut y avoir parthénogenèse ou apoméiose. Chez beaucoup de Chordariales enfin, la génération macroscopique de grande taille, morphologiquement évoluée, ne se développe qu'à une saison de l'année (l'été en général). L'espèce survit pendant l'hiver sous forme de générations microscopiques asexuées successives (microthalles ou pléthysmothalles). Ces individus microscopiques se multiplient abondamment par des zoïdes qui, dans certains cas tout au moins, peuvent être considérés comme des gamètes parthénogénétiques. Il semble qu'outre une perte fréquente de la sexualité il existe chez les Phéosporées une certaine indétermination dans la nature sexuée des cellules reproductrices.
Chez les Cyclosporées, il n'existe que des individus diploïdes, la méiose s'effectuant lors de la gamétogenèse, et le zygote se développe directement en un nouvel individu diploïde. Cela a conduit à considérer que les individus sont des gamétophytes diploïdes et que le cycle est monogénétique diplophasique.
Toutefois, certains auteurs proposent une interprétation différente. Selon celle-ci les individus diploïdes représenteraient un sporophyte dont les spores ne s'individualiseraient pas ; le gamétophyte auquel elles donnent normalement naissance chez les autres Phéophycées serait ici inclus dans le sporophyte (cas comparable à celui du prothalle femelle des Phanérogames), n'étant plus représenté chez la plupart des Cyclosporées que par la lignée cellulaire conduisant de la méiose à l'achèvement des gamètes.
Les Cyclosporées posséderaient donc, comme les autres Phéophycées, un cycle digénétique haplodiplophasique, mais chez elles le gamétophyte serait réduit et inclus dans le sporophyte diploïde, seul apparent. L'étude nouvelle de Cyclosporées primitives et de formes (Ascoseira) chez lesquelles la gamétogenèse s'effectue selon un processus moins simplifié que celui des Fucus trop souvent pris comme seul exemple, donne beaucoup de poids à cette hypothèse.
Accédez à l'intégralité de nos articles
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Jean FELDMANN : professeur à la faculté des sciences de Paris
Classification
Médias
Caractéristiques cytologiques
Encyclopædia Universalis France
Structures et types de croissance
Encyclopædia Universalis France
Ectocarpus : cycles de reproduction
Encyclopædia Universalis France
Autres références
-
ALGUES
- Écrit par Bruno DE REVIERS
- 4 869 mots
- 9 médias
 ...large) et d'organismes autotrophes comme les algues brun-doré (ou Ochrophyta, regroupant une quinzaine de classes d'algues dont les plus connues sont les algues brunes et les diatomées). Les alvéolés (A du groupe SAR) sont caractérisés par la présence d'alvéoles sous leur membrane cellulaire. Ils renferment...
...large) et d'organismes autotrophes comme les algues brun-doré (ou Ochrophyta, regroupant une quinzaine de classes d'algues dont les plus connues sont les algues brunes et les diatomées). Les alvéolés (A du groupe SAR) sont caractérisés par la présence d'alvéoles sous leur membrane cellulaire. Ils renferment... -
AQUACULTURE
- Écrit par Lucien LAUBIER
- 11 442 mots
- 8 médias
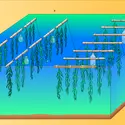 Commetoutes les Laminariales, l'algue brune Laminaria japonica (fig. 5) présente un cycle de vie digénétique, c'est-à-dire avec la succession d'une génération sporophytique (produisant des spores) très développée – pouvant atteindre plusieurs mètres – et une génération gamétophytique microscopique...
Commetoutes les Laminariales, l'algue brune Laminaria japonica (fig. 5) présente un cycle de vie digénétique, c'est-à-dire avec la succession d'une génération sporophytique (produisant des spores) très développée – pouvant atteindre plusieurs mètres – et une génération gamétophytique microscopique... -
DÉVELOPPEMENT, biologie
- Écrit par Georges DUCREUX , Hervé LE GUYADER et Jean-Claude ROLAND
- 19 226 mots
- 14 médias
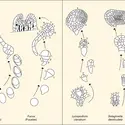 Les premières tentatives de développement embryonnaire apparaissent chez les algues. On peut en trouver des exemples chez lesPhéophycées – ou algues brunes – qui possèdent une grande diversité morphologique depuis des filaments à croissance indéterminée jusqu'à des thalles de grande taille –...
Les premières tentatives de développement embryonnaire apparaissent chez les algues. On peut en trouver des exemples chez lesPhéophycées – ou algues brunes – qui possèdent une grande diversité morphologique depuis des filaments à croissance indéterminée jusqu'à des thalles de grande taille –... -
GAMÈTES
- Écrit par Michel FAVRE-DUCHARTRE et Jacques TESTART
- 4 174 mots
- 5 médias
 Les gamètes hétérochontés, c'est-à-dire à flagelles inégaux, caractérisent les Chromophycophytes, surtout représentés par lesAlgues brunes (Fucus, par exemple). Originellement, l'un des flagelles est long et plumeux, l'autre court et lisse, mais la différence entre les deux flagelles peut...
Les gamètes hétérochontés, c'est-à-dire à flagelles inégaux, caractérisent les Chromophycophytes, surtout représentés par lesAlgues brunes (Fucus, par exemple). Originellement, l'un des flagelles est long et plumeux, l'autre court et lisse, mais la différence entre les deux flagelles peut... - Afficher les 7 références
Voir aussi
