- 1. Les étapes de la découverte
- 2. L'appareil photosynthétique
- 3. Les mécanismes primaires
- 4. Les réactions sombres : la réduction du dioxyde de carbone
- 5. Les types métaboliques de photosynthèse, d'après le mode de fixation initial de CO2
- 6. Photosynthèse nette et photosynthèse brute ; la photorespiration
- 7. Énergétique et importance de la photosynthèse dans la biosphère
- 8. Photosynthèse en C4
- 9. Bibliographie
PHOTOSYNTHÈSE
Article modifié le
L'appareil photosynthétique
Dans les feuilles vertes des plantes, dans les algues, la photosynthèse est réalisée par des organites spécialisés : les chloroplastes. Ils possèdent toujours une enveloppe formée de deux membranes : une externe perforée et une interne dotée de perméabilité sélective.
On y reconnaît la présence d'une structure fondamentale, la lamelle, remarquablement constante dans son architecture moléculaire générale. Il s'agit d'une bicouche lipidique (7 nm environ d'épaisseur), dans laquelle sont ancrées des macromolécules lipoprotéiques ou des complexes protéiques oligomériques. La fluidité des lipides membranaires (composés principalement de phospholipides et de galactolipides) autorise une certaine liberté de mouvement latéral de ces complexes. Toutes les molécules fonctionnellement associées aux étapes primaires de la photosynthèse sont intégrées, soit au sein de la membrane ( plastoquinones), soit comme protéines membranaires (cytochromes), soit liées de façon non covalente à celles-ci (chlorophylle des « antennes » et des « centres »). Mais certaines protéines [métalloprotéines telles la plastocyanine (Cu) ou la ferrédoxine (Fe)] s'associent de façon non permanente à certains sites de la membrane. Les plus gros complexes membranaires sont à la fois visibles en microscopie électronique et isolables après destruction ménagée de la membrane elle-même, de sorte qu'une localisation et une distribution assez précise de ces particules dans le plan de la membrane et selon sa normale a pu être abordée.

Chez les eucaryotes photosynthétiques (tous les végétaux chlorophylliens), il existe deux autres niveaux d'organisation englobant l'organisation supramacromoléculaire considérée ci-dessus. D'une part, les lamelles forment au sein du chloroplaste un réseau de vésicules ( thylakoïdes) qui définissent deux phases sans communication immédiate : l'espace stromatique et l'espace intrathylakoïdal (ou lumen). À la polarité stroma-lumen ainsi définie correspond une organisation transversale (normale au plan membranaire) des complexes et composants membranaires, qui est de la plus haute importance pour l'énergétique de la conversion (cf. fig. 7). D'autre part, les lamelles peuvent s'accoler par endroits pour former des empilements de disques ( granums, visibles en microscopie photonique) qui restent en relation de continuité avec des lamelles non accolées dispersées dans le stroma (fig. 3). On pense que cette disposition – fréquente mais non générale – représente une adaptation pour une répartition optimale de l'énergie lumineuse dans l'appareil.

Transport des électrons et des protons dans une membrane
Encyclopædia Universalis France
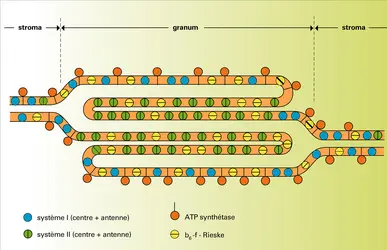
Thylakoïdes
Encyclopædia Universalis France
Il est relativement aisé de séparer, après broyage du tissu foliaire, des fragments de chloroplastes capables à la lumière de dégager de l'oxygène et de réduire certains accepteurs d'électrons (par exemple le ferricyanure et de nombreux colorants). Cette réaction de Hill, du nom du biochimiste anglais qui l'a découverte, permet de démontrer que c'est au sein des structures thylakoïdales que s'effectue l'intégralité de la conversion au sens énergétique. En effet, ces fragments de membranes isolés sont capables de réduire le NADP (nicotinamide-dinucléotide-phosphate) et de phosphoryler l'ADP (adénosine-diphosphate), à partir de quoi l'intégration du carbone en glucides peut s'effectuer de façon enzymatique spontanée.
On sait aller beaucoup plus loin dans la fragmentation de l'appareil en blocs élémentaires fonctionnels, notamment grâce à une attaque ménagée des membranes par des détersifs. Les centres réactionnels bactériens – où s'effectue l'acte photochimique de la conversion – donnent un bon exemple de cette démarche. On a pu déterminer la composition minimale d'un complexe, nécessaire à sa fonction : 3 sous-unités polypeptidiques, 4 molécules de bactériochlorophylle, 2 molécules de bactériophéophytine (macrocycle de chlorophylle privé de Mg), 1 molécule de caroténoïde, 2 molécules d'ubiquinone et 1 atome de fer. D'autres blocs fonctionnels sont difficiles à extraire et à purifier (stœchiométrie variable) ou perdent une partie de leur activité au cours de l'isolement. Mais le démontage de la micromachine photosynthétique découvre une remarquable unité d'architecture moléculaire – preuve évidente d'une parenté phylogénique – et nous apprend que la structure de ces complexes, c'est-à-dire la disposition géométrique de ses parties, tout autant que leur nature biochimique, détermine spécifiquement leurs fonctions.
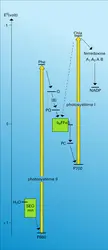
Schéma en Z des plantes supérieures
Encyclopædia Universalis France
La structure granaire – dominante chez les végétaux supérieurs – a longtemps posé un problème d'interprétation. À quoi sert-elle ? De nombreuses recherches ont été consacrées à ce problème. On s'est aperçu finalement que cette structure n'était pas statique, mais relativement mobile : sous l'influence de multiples facteurs (par exemple les ions Mg++ dans le stroma), les lamelles peuvent s'accoler ou se séparer réversiblement ; parallèlement, on assiste à une migration des complexes membranaires qui peuvent se rassembler ou se disperser. Un des résultats inattendus de ces recherches en un sens contredit le schéma en « Z », pourtant si solidement établi (fig. 5). Celui-ci, en effet, suggère naturellement qu'il doit exister dans la membrane une entité morphologique correspondant à la « chaîne photosynthétique » et constituée des deux principaux complexes : système II + système I. Or, si cette chaîne est bien une réalité sur le plan fonctionnel, son existence morphologique est extrêmement fluctuante. Ainsi, on a démontré, en les isolant, que les disques granaires ne comportent que des complexes système II et que tous les complexes système I (plus une minorité de système II) se rassemblent dans les lamelles dispersées dans le stroma. Il faut évidemment que des transporteurs mobiles – c'est le cas des plastoquinones – puissent établir une communication entre ces deux sortes d'édifices lamellaires. Cette singulière complication doit jouer un rôle, pense-t-on, dans l'adaptation de l'appareil photosynthétique au régime lumineux.
Accédez à l'intégralité de nos articles
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Jean LAVOREL : directeur de recherche honoraire au C.N.R.S., correspondant de l'Académie des sciences de Paris
- Paul MAZLIAK : professeur honoraire de biologie cellulaire, université de Paris-VI-Pierre-et-Marie-Curie
- Alexis MOYSE : professeur honoraire à l'université de Paris-Sud, correspondant de l'Académie des sciences
Classification
Médias

Chloroplastes et photosynthèse
Planeta Actimedia S.A.© Encyclopædia Universalis France pour la version française.
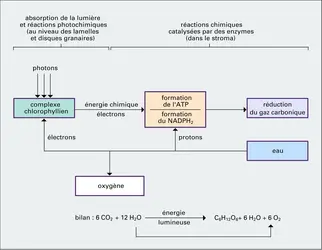
Réactions photochimiques et sombres
Encyclopædia Universalis France

Algue verte : spectres d'absorption et d'action
Encyclopædia Universalis France
Autres références
-
PHOTOSYNTHÈSE : CAPTURE DE L'ÉNERGIE LUMINEUSE
- Écrit par Claude LANCE
- 229 mots
Les travaux de Robert Emerson (1903-1959) ont permis d'élucider les mécanismes mis en jeu dans la photosynthèse notamment lors de la capture de l'énergie lumineuse. En 1932, Emerson confirme, avec W. Arnold, l'existence de deux phases distinctes dans ce processus : une phase photochimique,...
-
PHOTOSYNTHÈSE ET ÉNERGIE LUMINEUSE - (repères chronologiques)
- Écrit par Claude LANCE
- 531 mots
1845 J. R. Mayer, ayant formulé la loi de conservation de l'énergie, suggère que les plantes transforment 1'énergie solaire en énergie chimique.
1905 F. F. Blackman, en étudiant l'étude des facteurs limitants de la photosynthèse, notamment la température, est le...
-
PHOTOSYNTHÈSE ET FIXATION DU CO2 - (repères chronologiques)
- Écrit par Claude LANCE
- 345 mots
1837 H. von Mohl signale la présence de grains d'amidon dans les chloroplastes.
1843 J. von Liebig propose que 1'incorporation du dioxyde de carbone (CO2) se traduit par une formation d'acides organiques, précédant celle des sucres.
1862 J. von Sachs montre que les feuilles...
-
PHOTOSYNTHÈSE : UTILISATION DU CO2
- Écrit par Claude LANCE
- 259 mots
En faisant absorber à des végétaux du gaz carbonique marqué au carbone 14 (14CO2), le biochimiste américain Melvin Calvin (1911-1997) met en évidence, en 1954, le composé – ou accepteur – qui capte le CO2 de l'air dans le processus de la photosynthèse : il s'agit du ribulose-bisphosphate...
-
AGROMÉTÉOROLOGIE
- Écrit par Emmanuel CHOISNEL et Emmanuel CLOPPET
- 6 628 mots
- 7 médias
 ...tempérées), par rapport aux plantes en C4 (plantes ayant pour la plupart une origine tropicale, comme le maïs, la canne à sucre, le sorgho ou le millet). En effet, les plantes en C3 (plantes qui, par photosynthèse, fixent le dioxyde de carbone [CO2]en formant un composé à trois atomes de carbone) auront la...
...tempérées), par rapport aux plantes en C4 (plantes ayant pour la plupart une origine tropicale, comme le maïs, la canne à sucre, le sorgho ou le millet). En effet, les plantes en C3 (plantes qui, par photosynthèse, fixent le dioxyde de carbone [CO2]en formant un composé à trois atomes de carbone) auront la... -
ALGUES
- Écrit par Bruno DE REVIERS
- 4 869 mots
- 9 médias
 ...nommées plantes terrestres et qui regroupent, au sens large, les mousses, les fougères et les plantes à graines –, les algues possèdent de la chlorophylle. Ce pigment vert permet aux algues et aux embryophytes de fabriquer (synthétiser) de la matière vivante à partir du dioxyde de carbone (CO2) et de l'énergie...
...nommées plantes terrestres et qui regroupent, au sens large, les mousses, les fougères et les plantes à graines –, les algues possèdent de la chlorophylle. Ce pigment vert permet aux algues et aux embryophytes de fabriquer (synthétiser) de la matière vivante à partir du dioxyde de carbone (CO2) et de l'énergie... -
AUTOTROPHIE & HÉTÉROTROPHIE
- Écrit par Alexis MOYSE
- 2 504 mots
- 2 médias
 Les euglènes, algues unicellulaires des mares, possèdent de la chlorophylle et, parphotosynthèse, assimilent le gaz carbonique. Cependant elles ne peuvent vivre sur des milieux purement minéraux. Quelques substances organiques leur sont indispensables : la vitamine B12, les constituants de...
Les euglènes, algues unicellulaires des mares, possèdent de la chlorophylle et, parphotosynthèse, assimilent le gaz carbonique. Cependant elles ne peuvent vivre sur des milieux purement minéraux. Quelques substances organiques leur sont indispensables : la vitamine B12, les constituants de... -
BACTÉRIES
- Écrit par Jean-Michel ALONSO , Jacques BEJOT et Patrick FORTERRE
- 11 055 mots
- 3 médias
 ...Les bactéries et cyanobactéries capables d'utiliser l'énergie lumineuse sont les bactéries phototrophes. Parmi ces bactéries, certaines réalisent leurs photosynthèses en employant des composés minéraux comme donateurs d'électrons ; elles sont dites photolithotrophes ; d'autres ne peuvent utiliser...
...Les bactéries et cyanobactéries capables d'utiliser l'énergie lumineuse sont les bactéries phototrophes. Parmi ces bactéries, certaines réalisent leurs photosynthèses en employant des composés minéraux comme donateurs d'électrons ; elles sont dites photolithotrophes ; d'autres ne peuvent utiliser... - Afficher les 45 références
Voir aussi
- FIGUIER DE BARBARIE
- RENDEMENT, physico-chimie
- ENZYMES
- TRANSPIRATION VÉGÉTALE
- ABSORPTION, physique
- POACÉES ou GRAMINÉES
- CYTOSOL
- ATP SYNTHÉTASE
- PROTÉINES FER-SOUFRE
- STROMA
- DÉSERTS VIE DANS LES
- STRUCTURE, biologie
- THYLAKOÏDES
- POTENTIEL D'OXYDORÉDUCTION
- VÉGÉTALE BIOLOGIE
- EAU OXYGÉNÉE (peroxyde d'hydrogène)
- CALVIN CYCLE DE
- ALANINE
- CHIMIOSMOTIQUE THÉORIE
- DÉCARBOXYLATION
- CARBONE 14
- CONVERSION PHOTOCHIMIQUE
- NICOTINAMIDE ADÉNINE DINUCLÉOTIDE PHOSPHATE (NADP)
- ASPARTATE
- MÉSOPHYLLE
- PYRUVATE
- EMERSON ROBERT (1903-1959)
- PROTÉINES MEMBRANAIRES
- LIPIDES MEMBRANAIRES
- CARBONE CYCLE DU
- MARQUAGE ISOTOPIQUE
- PYRUVIQUE ACIDE
- RIBULOSE
- FRUCTOSE
- AMIDON
- PHYTOTECHNIE
- RÉDUCTION, chimie
- BIOSYNTHÈSES
- OXYDORÉDUCTIONS, biologie
- BIOÉNERGÉTIQUE
- OXALOACÉTATE
- MALATE
- ADAPTATION BIOLOGIQUE
- RESPIRATION
- PEROXYSOME
-
PHOTOSYNTHÈSE EN C
4 - TRANSPORT D'ÉLECTRONS, bioénergétique
- COUPLAGE, biochimie
- PLASTOQUINONE
- PHYSIOLOGIE VÉGÉTALE ou PHYTOBIOLOGIE
- GLYCOLATE OXYDASE
- RIBULOSE BISPHOSPHATE CARBOXYLASE
- PHOSPHOÉNOLPYRUVATE (PEP)
- FERRÉDOXINE
- MÉTABOLISME ACIDE CRASSULACÉEN ou CAM (Crassulacean Acid Metabolism)
- CARBONE 13
- GRANUM
- ÉCHANGES GAZEUX, physiologie
- ASSIMILATION, physiologie
- SPECTRE D'ABSORPTION
- CARBONIQUE GAZ ou DIOXYDE DE CARBONE
- SCIENCES HISTOIRE DES
