- 1. Les étapes de la découverte
- 2. L'appareil photosynthétique
- 3. Les mécanismes primaires
- 4. Les réactions sombres : la réduction du dioxyde de carbone
- 5. Les types métaboliques de photosynthèse, d'après le mode de fixation initial de CO2
- 6. Photosynthèse nette et photosynthèse brute ; la photorespiration
- 7. Énergétique et importance de la photosynthèse dans la biosphère
- 8. Photosynthèse en C4
- 9. Bibliographie
PHOTOSYNTHÈSE
Article modifié le
Les types métaboliques de photosynthèse, d'après le mode de fixation initial de CO2
Les réactions enzymatiques décrites ci-dessus sont sensiblement les seules voies d'entrée du CO2 dans le métabolisme photosynthétique, pour la majorité des plantes primitivement étudiées. 90 p. 100 du carbone au moins empruntent directement la voie du phosphoglycérate, composé à 3 atomes de C, d'où le qualificatif de « type C3 » donné à ces végétaux qui comprennent presque toutes les plantes originaires des régions tempérées, toutes les fougères, mousses, algues et enfin tous les arbres quel que soit leur habitat climatique.
Mais, parmi les plantes herbacées et quelques arbustes des régions tropicales, semi-tropicales, ou encore des sols salés des régions tempérées, on trouve, dans près de vingt familles appartenant toutes aux phanérogames angiospermes, un procédé différent de fixation initial de CO2. Celui-ci a été reconnu entre 1963 et 1966. Dans le cytosol des cellules sous-épidermiques de leurs feuilles ( mésophylle) la réaction suivante intervient pour la quasi-totalité du CO2 fixé. Elle est catalysée par une phosphoénolpyruvate-carboxylase :

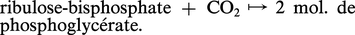
Cycles des métabolites
Encyclopædia Universalis France
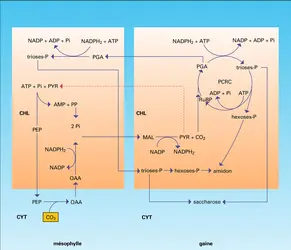
Cette dernière a lieu dans les chloroplastes exclusivement. Ces cellules, plus internes que celles du mésophylle, entourent les vaisseaux conducteurs de sève et constituent la gaine périvasculaire (fig. 11). L'ensemble forme une structure dite en « couronne ». Puis les opérations de réduction du PGA et de transformation des glucides, avec régénération du ribulose-diphosphate, formation d'amidon et de saccharose, se poursuivent comme dans les plantes de type C3. La figure 12 schématise les 2 cycles réalisés, l'un dans les cellules du mésophylle, l'autre dans celles de la gaine périvasculaire. Ainsi l'ensemble du métabolisme photosynthétique du carbone se trouve réparti entre deux tissus dont les cellules ont une compartimentation enzymatique nettement diversifiée.

De plus, le type C4 présente plusieurs variantes. Pour le maïs, la canne à sucre, le malate est le transporteur essentiel de CO2 et, dans les chloroplastes des cellules des gaines, une enzyme malique catalyse la réaction :
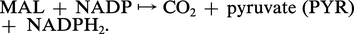
Le malate fournit le CO2 et enrichit également le pouvoir réducteur des chloroplastes des gaines en réduisant le NADP en NADPH2. Pour d'autres espèces de type C4 : Panicum maximum, Chloris guyana, l'aspartate, après amination de l' oxaloacétate dans le cytoplasme des cellules du mésophylle, émigre dans les gaines, y régénère l'OAA qui enfin fournit le CO2 aux chloroplastes de ces dernières, après une décarboxylation par une PEP-carboxykinase selon une réaction du type :
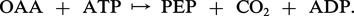
Le phosphoénolpyruvate, à l'état de pyruvate, est ensuite aminé, donnant naissance à l'α-alanine qui retourne dans le mésophylle, assurant la fourniture à la fois d'un chaînon tricarboné et d'une fonction aminée nécessaire à la poursuite du cycle.
Pour les espèces telles que Amaranthus edulis, Panicum miliaceum, où l'aspartate est également la principale forme de transport de CO2 du mésophylle aux cellules des gaines, cet acide aminé est reconverti en malate dans les mitochondries des cellules des gaines et une enzyme malique assure la libération du CO2 selon la réaction :

Après amination du pyruvate, avec formation d'α-alanine, le retour de cet acide aminé tricarboné permet la reprise du cycle dans les cellules du mésophylle.
Si, dans ces trois modalités, le cycle réducteur du CO2 en glucides se situe toujours dans les chloroplastes des cellules des gaines périvasculaires et par le même processus incluant la carboxylation du ribulose-diphosphate avec formation de PGA et réduction de ce dernier en trioses-phosphates, le mode d'apport de CO2 à l'intérieur des tissus présente des modalités différentes. Le métabolisme photosynthétique du carbone est « éclaté », partagé entre des lieux cellulaires ou des organites divers : cytosol, chloroplastes, voire mitochondries, avec une spécialisation des cellules sous-épidermiques externes du mésophylle dans la fixation initiale du CO2 et une spécialisation des chloroplastes des cellules plus interne des gaines dans sa réduction au niveau glucidique. Il en résulte une compartimentation du métabolisme différente de celle des plantes de type C3.
D'autres complications interviennent d'ailleurs, les chloroplastes des cellules du mésophylle pouvant participer à la réduction d'une fraction du phosphoglycérate venant des plastes des gaines, mais seuls les chloroplastes de ces dernières possèdent la ribulose-diphosphate-carboxylase.
Cet éclatement avec une compartimentation complexe implique diverses migrations de métabolites intermédiaires, facilitées par des communications directes (plasmodesmes entre les cytosols des différentes cellules en continuité, qu'elles appartiennent au mésophylle ou aux gaines). Cette compartimentation implique aussi une dépense énergétique supplémentaire correspondant à la multiplicité des transports et aux réactions plus nombreuses quand on compare le métabolisme C4 au métabolisme C3. Mais cette dépense supplémentaire est couverte par l'adaptation des plantes de type C4 au fort ensoleillement de leurs régions d'élection. Par ailleurs, ainsi qu'on l'exposera plus loin, à propos de la photorespiration, le pouvoir de fixation de CO2 de la PEP-carboxylase est beaucoup plus élevé que celui de la ribulose-diphosphate-carboxylase et leur assure une beaucoup plus grande activité métabolique de photosynthèse quand le flux lumineux n'est pas limitant.
Les plantes de type C4 sont capables d'absorber la totalité du CO2 présent dans une enceinte close, ce que ne peuvent réaliser les plantes de type C3 dont la photosynthèse s'annule lorsque la pression partielle de CO2 dans l'atmosphère s'abaisse au-dessous de 40 à 50 microlitres par litre d'air (point de compensation de CO2).
De ce fait, en atmosphère normale, les plantes de type C4 absorbent presque aussi rapidement le CO2 dont le carbone est l'isotope 13C que le CO2 à carbone 12C (le 13CO2 représente environ 1,1 p. 100 du CO2 total de l'air), alors que les plantes de type C3 présentent à l'égard du 13CO2 un retard à l'assimilation de 20 p. 1 000 par rapport au 12CO2 (fractionnement isotopique), contre 4 p. 1000 pour les plantes de type C4.
Elles peuvent utiliser des éclairements beaucoup plus élevés que les plantes de type C3. Leur saturation lumineuse n'est pas toujours atteinte dans les jours les plus riches en lumière (500 W par m2), alors que les plantes de type C3 sont saturées de lumière entre 50 et 150 watts par mètre carré. Leur optimum thermique de photosynthèse est également plus élevé (de 30 à 47 0C contre 15 à 25 0C pour les plantes de type C3).
La structure en couronne des tissus foliaires des plantes de type C4 est favorable à la migration des produits de photosynthèse, en raison de la proximité des cellules des gaines où se réalise la synthèse des glucides (amidon et saccharose) et des vaisseaux conducteurs de la sève qui distribuent les métabolites aux divers organes de la plante et peuvent ainsi leur assurer une croissance plus rapide, dans les conditions climatiques qui leur sont favorables.
Un autre avantage leur est aussi conféré dans leurs régions d'origine par leur économie hydrique, leur transpiration étant, en présence d'un ensoleillement fort, plus faible que celui des plantes de type C3 et leur rapport carbone assimilé/eau transitant dans les tissus plus élevé. Elles se trouvent sur la voie d'une adaptation aux climats arides dont le terme est donné par les plantes grasses au métabolisme acide crassulacéen (CAM) qui constitue un troisième type adapté aux conditions des déserts aux températures diurnes élevées. Au moins vingt-six familles de phanérogames et quelques fougères possèdent des espèces de type CAM.

Figuier de Barbarie
unjiko/ Shutterstock
En 1804, N. T. de Saussure avait décrit que les raquettes de figuier-de-Barbarie (Opuntia ficus-indica) fixaient le CO2 la nuit, quand les orifices de leurs stomates sont ouverts. Le jour, ils sont fermés et la transpiration de la plante est ainsi supprimée pendant la période de chaleur sèche de leur climat d'origine. La fixation aboutit alors, comme dans les plantes de type C4, à la formation de malate qui s'accumule durant la nuit, dans les vacuoles des cellules. Le jour suivant, à la lumière, le malate fournit le CO2 nécessaire à la synthèse des glucides qui a lieu dans les chloroplastes. Il y a disjonction dans le temps entre la fixation initiale de CO2 et sa réduction.
Il existe des espèces végétales dont les caractères sont intermédiaires entre les types métaboliques précédemment décrits, mais elles sont apparemment moins fréquentes que les types tranchés. Sans doute témoignent-elles des modalités d'évolution avortées ou en cours. Le type C3 est le plus primitif des trois, d'après les fossiles connus et les caractéristiques de la composition isotopique en carbone 13 et en carbone 12 des charbons. Il faut noter aussi que dans la même famille botanique (Caryophyllaceae, Chenopodiaceae, Euphorbiaceae...) se trouvent des représentants des trois types métaboliques.
Enfin, des essais d'hybridation ont été tentés entre espèces voisines de type C3 ou de type C4 appartenant au même genre (Atriplex). Les croisements donnent des descendances viables, mais les nombreux caractères distinctifs, anatomiques, cytologiques, enzymatiques, dépendent de gènes différents. Ils se disjoignent avec une forte indépendance, une grande irrégularité dans la distribution chromosomique, qui, jusqu'à présent, n'a pu être maîtrisée en vue de conduire à une amélioration comparable à celle des plantes de grande culture telles que les blés ou les maïs.
Accédez à l'intégralité de nos articles
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Jean LAVOREL : directeur de recherche honoraire au C.N.R.S., correspondant de l'Académie des sciences de Paris
- Paul MAZLIAK : professeur honoraire de biologie cellulaire, université de Paris-VI-Pierre-et-Marie-Curie
- Alexis MOYSE : professeur honoraire à l'université de Paris-Sud, correspondant de l'Académie des sciences
Classification
Médias

Chloroplastes et photosynthèse
Planeta Actimedia S.A.© Encyclopædia Universalis France pour la version française.
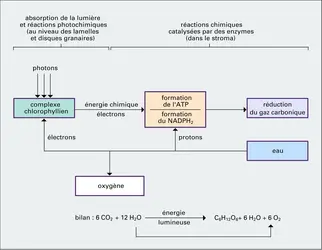
Réactions photochimiques et sombres
Encyclopædia Universalis France

Algue verte : spectres d'absorption et d'action
Encyclopædia Universalis France
Autres références
-
PHOTOSYNTHÈSE : CAPTURE DE L'ÉNERGIE LUMINEUSE
- Écrit par Claude LANCE
- 229 mots
Les travaux de Robert Emerson (1903-1959) ont permis d'élucider les mécanismes mis en jeu dans la photosynthèse notamment lors de la capture de l'énergie lumineuse. En 1932, Emerson confirme, avec W. Arnold, l'existence de deux phases distinctes dans ce processus : une phase photochimique,...
-
PHOTOSYNTHÈSE ET ÉNERGIE LUMINEUSE - (repères chronologiques)
- Écrit par Claude LANCE
- 531 mots
1845 J. R. Mayer, ayant formulé la loi de conservation de l'énergie, suggère que les plantes transforment 1'énergie solaire en énergie chimique.
1905 F. F. Blackman, en étudiant l'étude des facteurs limitants de la photosynthèse, notamment la température, est le...
-
PHOTOSYNTHÈSE ET FIXATION DU CO2 - (repères chronologiques)
- Écrit par Claude LANCE
- 345 mots
1837 H. von Mohl signale la présence de grains d'amidon dans les chloroplastes.
1843 J. von Liebig propose que 1'incorporation du dioxyde de carbone (CO2) se traduit par une formation d'acides organiques, précédant celle des sucres.
1862 J. von Sachs montre que les feuilles...
-
PHOTOSYNTHÈSE : UTILISATION DU CO2
- Écrit par Claude LANCE
- 259 mots
En faisant absorber à des végétaux du gaz carbonique marqué au carbone 14 (14CO2), le biochimiste américain Melvin Calvin (1911-1997) met en évidence, en 1954, le composé – ou accepteur – qui capte le CO2 de l'air dans le processus de la photosynthèse : il s'agit du ribulose-bisphosphate...
-
AGROMÉTÉOROLOGIE
- Écrit par Emmanuel CHOISNEL et Emmanuel CLOPPET
- 6 628 mots
- 7 médias
 ...tempérées), par rapport aux plantes en C4 (plantes ayant pour la plupart une origine tropicale, comme le maïs, la canne à sucre, le sorgho ou le millet). En effet, les plantes en C3 (plantes qui, par photosynthèse, fixent le dioxyde de carbone [CO2]en formant un composé à trois atomes de carbone) auront la...
...tempérées), par rapport aux plantes en C4 (plantes ayant pour la plupart une origine tropicale, comme le maïs, la canne à sucre, le sorgho ou le millet). En effet, les plantes en C3 (plantes qui, par photosynthèse, fixent le dioxyde de carbone [CO2]en formant un composé à trois atomes de carbone) auront la... -
ALGUES
- Écrit par Bruno DE REVIERS
- 4 869 mots
- 9 médias
 ...nommées plantes terrestres et qui regroupent, au sens large, les mousses, les fougères et les plantes à graines –, les algues possèdent de la chlorophylle. Ce pigment vert permet aux algues et aux embryophytes de fabriquer (synthétiser) de la matière vivante à partir du dioxyde de carbone (CO2) et de l'énergie...
...nommées plantes terrestres et qui regroupent, au sens large, les mousses, les fougères et les plantes à graines –, les algues possèdent de la chlorophylle. Ce pigment vert permet aux algues et aux embryophytes de fabriquer (synthétiser) de la matière vivante à partir du dioxyde de carbone (CO2) et de l'énergie... -
AUTOTROPHIE & HÉTÉROTROPHIE
- Écrit par Alexis MOYSE
- 2 504 mots
- 2 médias
 Les euglènes, algues unicellulaires des mares, possèdent de la chlorophylle et, parphotosynthèse, assimilent le gaz carbonique. Cependant elles ne peuvent vivre sur des milieux purement minéraux. Quelques substances organiques leur sont indispensables : la vitamine B12, les constituants de...
Les euglènes, algues unicellulaires des mares, possèdent de la chlorophylle et, parphotosynthèse, assimilent le gaz carbonique. Cependant elles ne peuvent vivre sur des milieux purement minéraux. Quelques substances organiques leur sont indispensables : la vitamine B12, les constituants de... -
BACTÉRIES
- Écrit par Jean-Michel ALONSO , Jacques BEJOT et Patrick FORTERRE
- 11 055 mots
- 3 médias
 ...Les bactéries et cyanobactéries capables d'utiliser l'énergie lumineuse sont les bactéries phototrophes. Parmi ces bactéries, certaines réalisent leurs photosynthèses en employant des composés minéraux comme donateurs d'électrons ; elles sont dites photolithotrophes ; d'autres ne peuvent utiliser...
...Les bactéries et cyanobactéries capables d'utiliser l'énergie lumineuse sont les bactéries phototrophes. Parmi ces bactéries, certaines réalisent leurs photosynthèses en employant des composés minéraux comme donateurs d'électrons ; elles sont dites photolithotrophes ; d'autres ne peuvent utiliser... - Afficher les 45 références
Voir aussi
- FIGUIER DE BARBARIE
- RENDEMENT, physico-chimie
- ENZYMES
- TRANSPIRATION VÉGÉTALE
- ABSORPTION, physique
- POACÉES ou GRAMINÉES
- CYTOSOL
- ATP SYNTHÉTASE
- PROTÉINES FER-SOUFRE
- STROMA
- DÉSERTS VIE DANS LES
- STRUCTURE, biologie
- THYLAKOÏDES
- POTENTIEL D'OXYDORÉDUCTION
- VÉGÉTALE BIOLOGIE
- EAU OXYGÉNÉE (peroxyde d'hydrogène)
- CALVIN CYCLE DE
- ALANINE
- CHIMIOSMOTIQUE THÉORIE
- DÉCARBOXYLATION
- CARBONE 14
- CONVERSION PHOTOCHIMIQUE
- NICOTINAMIDE ADÉNINE DINUCLÉOTIDE PHOSPHATE (NADP)
- ASPARTATE
- MÉSOPHYLLE
- PYRUVATE
- EMERSON ROBERT (1903-1959)
- PROTÉINES MEMBRANAIRES
- LIPIDES MEMBRANAIRES
- CARBONE CYCLE DU
- MARQUAGE ISOTOPIQUE
- PYRUVIQUE ACIDE
- RIBULOSE
- FRUCTOSE
- AMIDON
- PHYTOTECHNIE
- RÉDUCTION, chimie
- BIOSYNTHÈSES
- OXYDORÉDUCTIONS, biologie
- BIOÉNERGÉTIQUE
- OXALOACÉTATE
- MALATE
- ADAPTATION BIOLOGIQUE
- RESPIRATION
- PEROXYSOME
-
PHOTOSYNTHÈSE EN C
4 - TRANSPORT D'ÉLECTRONS, bioénergétique
- COUPLAGE, biochimie
- PLASTOQUINONE
- PHYSIOLOGIE VÉGÉTALE ou PHYTOBIOLOGIE
- GLYCOLATE OXYDASE
- RIBULOSE BISPHOSPHATE CARBOXYLASE
- PHOSPHOÉNOLPYRUVATE (PEP)
- FERRÉDOXINE
- MÉTABOLISME ACIDE CRASSULACÉEN ou CAM (Crassulacean Acid Metabolism)
- CARBONE 13
- GRANUM
- ÉCHANGES GAZEUX, physiologie
- ASSIMILATION, physiologie
- SPECTRE D'ABSORPTION
- CARBONIQUE GAZ ou DIOXYDE DE CARBONE
- SCIENCES HISTOIRE DES
