- 1. La disparité des métazoaires : plans d'organisation et embranchements
- 2. La conception traditionnelle de l'évolution animale
- 3. Le renouveau des études
- 4. La monophylie des métazoaires et leur position dans l’arbre des eucaryotes
- 5. La base de l'arbre : un problème non résolu
- 6. La phylogénie et l'évolution des Bilateria
- 7. La contribution des données paléontologiques
- 8. La contribution de l'évo-dévo
- 9. Vers un arbre entièrement résolu des métazoaires ?
- 10. Bibliographie
PHYLOGÉNIE ANIMALE
Article modifié le
La contribution des données paléontologiques
Les fossiles susceptibles d'apporter des informations sur les relations entre les embranchements actuels et sur l’origine des plans d'organisation sont essentiellement ceux qui sont compris entre 600 et 500 millions d'années (Ma), ce qui correspond à la fin du Protérozoïque et au Cambrien. Leur exploitation est toutefois rendue délicate du fait de l'existence de lacunes et de la difficulté que pose l'interprétation de certaines traces fossiles.
Certaines formations de la fin du Protérozoïque (étage Vendien) renferment de nombreuses empreintes d'animaux à corps mou qui sont communément interprétés comme des éponges, des cnidaires, des Bilateria primitifs (par exemple Spriggina) et des formes encore énigmatiques. Cette fameuse faune d'Ediacara a d'abord été découverte en Australie, dans les monts Ediacara (d'où son nom), puis retrouvée sur tous les continents. Datée de 570 à 550 Ma, elle résulterait de la première radiation des métazoaires, ayant produit les premières branches de l'arbre des métazoaires (fig. 3). Certains chercheurs proposent des interprétations alternatives : par exemple, pour Seilacher, la plupart des fossiles d'Ediacara seraient des organismes photosynthétiques syncytiaux sans aucun rapport avec des métazoaires.
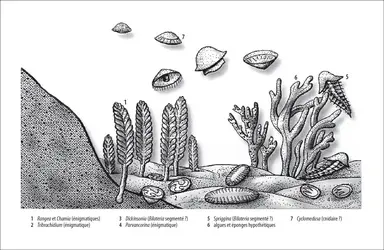
Faune d'Ediacara
Encyclopædia Universalis France
Le début du Cambrien, aujourd'hui daté à 543 Ma, se caractérise par une augmentation considérable des traces de bioturbation dans les sédiments, par l'apparition des terriers verticaux (ce qui indique la présence d'animaux capables de locomotion fouisseuse active) et des structures squelettiques minéralisées (en fait, les tout premiers tubes calcaires, Cloudina, s'observent dès l'extrême fin du Protérozoïque). Par ailleurs, dès le Cambrien inférieur apparaissent les embranchements modernes de Bilateria (mollusques, arthropodes, vertébrés, etc.). Au Cambrien moyen, pratiquement tous les embranchements actuels des Bilateria sont présents dans les riches faunes de Burgess (530-520 Ma) [fig. 6]. Leur apparition quasi simultanée à l'échelle des temps géologiques rend impossible l'établissement d'une chronologie. Ainsi, selon l'interprétation classique, les embranchements des Bilateria seraient issus d'une radiation évolutive extrêmement rapide (à l'échelle des temps géologiques), que l'on a appelée l'« explosion cambrienne ».
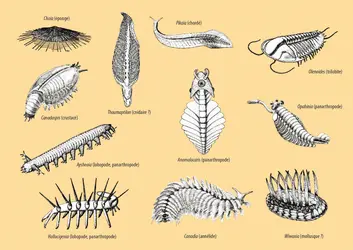
Faune de Burgess
Encyclopædia Universalis France
Le paléontologue et évolutionniste américain Stephen Jay Gould (1941-2002) a largement popularisé ces fossiles des schistes de Burgess et leur importance du point de vue de l'évolution animale dans son ouvrage Wonderful Life, publié en 1989 (La vie est belle, 1994). L'une des idées maîtresses de ce livre est que la disparité anatomique de cette faune dépassait celle du temps présent. Un certain nombre de plans d'organisation auraient disparu par la suite, non pas, selon Gould, parce qu'ils étaient moins performants, mais simplement par hasard (du fait notamment des aléas environnementaux). Il faut toutefois mentionner que, depuis lors, pratiquement tous les fossiles « énigmatiques » des faunes de Burgess ont pu être reclassés dans ou près d'embranchements existant de nos jours (par exemple Hallucigenia, réinterprété comme un panarthropode après avoir été retourné dorso-ventralement, ou encore Odontogriphus, récemment réinterprété comme un mollusque). En particulier, les nombreux fossiles du Cambrien apparentés aux panarthropodes actuels, ou encore aux échinodermes actuels, s'avèrent extrêmement utiles pour clarifier l'évolution des structures anatomiques dans ces régions de l’arbre.

Ces dernières années, de nombreuses études ont cherché à dater les nœuds profonds de l'arbre des métazoaires en utilisant le degré de divergence des séquences de gènes (datation moléculaire). Ainsi, les dates obtenues pour la divergence entre protostomiens et deutérostomiens (donc pour l'ancêtre le plus récent de tous les Bilateria) varient dans une fourchette de 1 000 à 600 Ma. Bien qu'imprécises, ces datations remettent en cause l'idée d'une radiation des Bilateria au début du Cambrien ; cette dernière serait alors plus ancienne que les faunes d'Ediacara.
Ces résultats ont suscité de nouvelles interprétations des données paléontologiques : ou bien la brusque apparition des embranchements de Bilateria au Cambrien serait simplement due à des conditions particulières de fossilisation et ne refléterait en rien l'origine de ces lignées, qui auraient une longue histoire non encore dévoilée au Protérozoïque ; ou bien l'explosion cambrienne correspondrait bien à l'apparition des embranchements modernes, mais par des radiations indépendantes au sein des trois grands clades (ecdysozoaires, lophotrochozoaires, deutérostomiens), la radiation ayant donné naissance à ces grands clades eux-mêmes étant plus ancienne. Dans ce cas aussi, les Bilateria auraient eu une histoire secrète plus ou moins longue au Protérozoïque.
Incités dans ce contexte à rechercher des Bilateria dans les formations protérozoïques, des paléontologues en ont mis au jour dans des gisements phosphoritiques de Chine (faune de Doushantuo, 600-580 Ma), surtout depuis les années 1990. Ces sédiments présentent une préservation exceptionnelle, permettant d'observer les détails de petites structures même à l'échelle cellulaire. Il s'agit par exemple de microfossiles interprétés comme des embryons de deutérostomiens (gastrula avec sacs cœlomiques rattachés à l'archentéron), ou encore du remarquable Vernanimalcula, dont l'anatomie comporterait un cœlome très net.
L'interprétation de ces fossiles est cependant très discutable. Des précipitations d'origine purement minérale peuvent, par exemple, doter une cavité d'un revêtement évoquant une paroi cœlomique. Un problème récurrent de la paléontologie animale du Protérozoïque et du début du Cambrien est que la nature zoologique des formes différant le plus des embranchements actuels (potentiellement les plus informatives) est en général obscure, par manque d'informations suffisamment détaillées sur l'anatomie.
Accédez à l'intégralité de nos articles
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Michaël MANUEL : ancien élève de l'École normale supérieure, agrégé des sciences de la vie et de la Terre, maître de conférences à l'université Paris-VI
Classification
Voir aussi
- FEUILLETS EMBRYONNAIRES ou GERMINATIFS
- MÉSODERME
- ECTODERME
- ENDODERME, embryologie animale
- CŒLOME
- GÈNES HOMÉOTIQUES
- VESTIMENTIFÈRES
- NÉMATODES
- CŒLOMATES
- PSEUDOCŒLOMATES
- SYSTÉMATIQUE
- DÉVELOPPEMENT ANIMAL ou ONTOGENÈSE ANIMALE
- DEUTÉROSTOMIENS
- PLAN D'ORGANISATION, zoologie
- ZOOLOGIE
- PROTOSTOMIENS
- ÉPONGE
- ECHIURIENS
- EMBRANCHEMENT, systématique
- MUE
- TRIPLOBLASTIQUES ANIMAUX
- MÉTAZOAIRES
- GRADE, biologie
- PARAPHYLIE, systématique
- ANIMAL RÈGNE
- PHYLOGÉNIE ou PHYLOGENÈSE
- ANATOMIE ANIMALE
- MÉTAMÈRE ou SEGMENT
- CŒLENTÉRÉS
- DIPLOBLASTIQUES ANIMAUX
- ACŒLOMATES
- CLADE
- SYNAPOMORPHIE
- PALÉOZOOLOGIE
- PHORONIDIENS
- POLYZOAIRES ou BRYOZOAIRES
- CONVERGENCE ÉVOLUTIVE
- CARACTÈRES, biologie
- MONOPHYLIE, systématique
- BIOLOGIE HISTOIRE DE LA
- BILATERIA
- RÉCAPITULATION THÉORIE DE LA ou LOI BIOGÉNÉTIQUE FONDAMENTALE
- PHYLOGÉNIE MOLÉCULAIRE
- CLADOGRAMME
- EUMÉTAZOAIRES
- TRICHOPLAX
- PLACOZOAIRES
- LOPHOTROCHOZOAIRES
- ECDYSOZOAIRES
- ACŒLES
- NÉMERTODERMATIDES
- ACŒLOMORPHES
- SCALIDOPHORES
- NÉMATOÏDES
- PANARTHROPODES
- AMBULACRALIA
- PHARYNGOTRÉMIE
- XENOTURBELLA
- SIBOGLINIDÉS
- DATATION MOLÉCULAIRE, biologie
- ÉVO-DÉVO (evolutionary developmental biology)
- URBILATERIA
- ANCÊTRE COMMUN, phylogénie
- POLYPHYLIE, systématique
- ARBRE PHYLOGÉNÉTIQUE
