
- 1. Histoire de la phytopathologie
- 2. Maladies des plantes et symptômes
- 3. Diversité des agents pathogènes
- 4. Virulence des agents pathogènes
- 5. Diversité des défenses des plantes
- 6. Réponses immunitaires des plantes
- 7. Coévolution virulence-défense et niveaux de résistance aux maladies
- 8. Comprendre pour protéger
- 9. Bibliographie
PHYTOPATHOLOGIE
Réponses immunitaires des plantes
Les réponses immunitaires des plantes sont déclenchées par la perception du « non-soi », laquelle peut être de type général ou plus spécifique selon la nature des molécules reconnues.
La réponse de type général est consécutive à la reconnaissance de motifs moléculaires communs à de nombreux microorganismes. Dénommés éliciteurs et plus récemment PAMP ou MAMP (pathogen/microbe associatedmolecular patterns), ces motifs, exogènes aux plantes, sont de nature chimique diverse (glucidique, protéique, lipidique). Ils proviennent essentiellement de polymères de structure des surfaces cellulaires tels que la chitine et les glucanes des champignons et des oomycètes, le lipopolysaccharide et la protéine flagelline des bactéries, ainsi que des éléments structuraux des virus. Plusieurs de ces motifs sont libérés sous l’action des enzymes hydrolytiques des plantes. La perception du « soi altéré », c’est-à-dire d’oligosaccharides résultant de la dégradation des parois cellulaires de l’hôte par les enzymes des agents pathogènes, fonctionne également comme signal d’alarme, induisant des réactions de défense. Ces oligosaccharides sont donc des éliciteurs endogènes, déclenchant les mécanismes de défense de la plante.
Les éliciteurs sont des outils puissants qui ont permis de décrypter aux niveaux cellulaire et moléculaire les cascades de signaux qu’ils génèrent à partir de leur reconnaissance par des protéines situées en surface des plantes. Appelées PRR (pattern recognition receptors), ces protéines renferment des motifs structuraux répétés riches en leucine (acide aminé) et impliqués dans les interactions avec les éliciteurs. Les signaux générés par cette interaction – de nature ionique (calcium, formes activées de l’oxygène), hormonale (éthylène, acide salicylique, acide jasmonique), protéique (kinases, facteurs de transcription), etc. – régulent à leur tour l’expression des gènes de défense évoqués précédemment. La réceptivité aux éliciteurs revêt un caractère général : quelle que soit sa provenance (microorganisme, plante), un éliciteur donné peut stimuler le système de défense de nombreuses plantes contre divers agents pathogènes. C’est donc la résistance de base aux maladies qui est déclenchée par les éliciteurs. Aussi appelée PTI (pour patterntriggeredimmunity), cette résistance est la composante non spécifique de la réponse immunitaire des plantes, un mécanisme pour partie commun aux plantes et aux animaux.
La mise en évidence d’une reconnaissance spécifique du « non-soi » découle de l’existence de couples hôte-parasite à l’intérieur desquels il existe des interactions différentielles hautement spécifiques entre différentes variétés (sensibles ou résistantes) d’une même espèce d’hôte et différentes races (virulentes ou avirulentes) d’une même espèce de parasite. Une variété donnée peut être résistante à une race donnée mais sensible à une autre race. C’est à partir d’une telle interaction entre le lin et le champignonMelampsoralini, responsable de la rouille, que Harold Flor a établi en 1946-1947 les bases génétiques de la spécificité. En étudiant la ségrégation des caractères conditionnant la maladie, il montre, d’une part, que la résistance de l’hôte et l’avirulence du parasite sont des caractères dominants et, d’autre part, que les phénotypes conférés par ces caractères sont portés par un seul gène côté plante et un seul gène côté agent pathogène. Ainsi, la présence d’un gène de résistance dominant (R) et d’un gène d’avirulence dominant (Avr) conduit à une interaction incompatible (R/Avr) : la maladie ne se développe pas, la plante est résistante. L’interaction compatible (la plante est sensible, il y a maladie) survient lorsque l’un de ces deux gènes partenaires ou les deux sont non fonctionnels[...]
La suite de cet article est accessible aux abonnés
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Marie-Thérèse ESQUERRÉ-TUGAYÉ : professeure émérite des Universités, membre de l'Académie d'agriculture de France
Classification
Médias

Mildiou
A. & H.-F. Michler/ Science Photo Library/ Biosphoto

Mosaïque du tabac
N. Cattlin/ Visuals Unlimited/ Corbis

Cloque du pêcher
canoniroff/ Shutterstock
Autres références
-
ACARIENS
- Écrit par Jean-Louis CONNAT et Gabriel GACHELIN
- 6 631 mots
- 2 médias
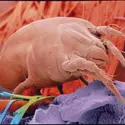 De nombreux acariens sont également des parasites de plantes. De très petite taille (longueur de 0,1 à 0,5 mm), ils causent des dégâts très variables. Le plus fréquent, pour les zones tempérées, est l'acarien rouge (genre Panonychus) des arbres fruitiers, qui affecte également les rosiers....
De nombreux acariens sont également des parasites de plantes. De très petite taille (longueur de 0,1 à 0,5 mm), ils causent des dégâts très variables. Le plus fréquent, pour les zones tempérées, est l'acarien rouge (genre Panonychus) des arbres fruitiers, qui affecte également les rosiers.... -
ACTINOMYCÈTES
- Écrit par Hubert A. LECHEVALIER
- 3 450 mots
- 4 médias
-
AGROMÉTÉOROLOGIE
- Écrit par Emmanuel CHOISNEL et Emmanuel CLOPPET
- 6 627 mots
- 7 médias
 Lesconditions météorologiques peuvent jouer un rôle prépondérant dans le déroulement d'une séquence épidémique. Le climat agit en effet à la fois sur la plante et sur l'agent pathogène (champignon, bactérie, virus, etc.). Ainsi la température conditionne en partie, d'une part, le déroulement du cycle...
Lesconditions météorologiques peuvent jouer un rôle prépondérant dans le déroulement d'une séquence épidémique. Le climat agit en effet à la fois sur la plante et sur l'agent pathogène (champignon, bactérie, virus, etc.). Ainsi la température conditionne en partie, d'une part, le déroulement du cycle... -
AGRONOMIE
- Écrit par Stéphane HÉNIN et Michel SEBILLOTTE
- 9 202 mots
- 1 média
 ...malédiction céleste ; mais l'homme s'est rapidement aperçu qu'au moins les causes secondes étaient liées à la présence de parasites animaux et végétaux. Le citadin imagine mal de nos jours ce qu'ont pu être ces fléaux, mais il peut s'en faire une idée en sachant que, vers 1840, l'apparition en Irlande...
...malédiction céleste ; mais l'homme s'est rapidement aperçu qu'au moins les causes secondes étaient liées à la présence de parasites animaux et végétaux. Le citadin imagine mal de nos jours ce qu'ont pu être ces fléaux, mais il peut s'en faire une idée en sachant que, vers 1840, l'apparition en Irlande... - Afficher les 25 références
