- 1. Diagnose
- 2. Classification
- 3. Catégories systématiques (groupes zoologiques)
- 4. Habitat et comportement alimentaire
- 5. Biologie sexuelle et reproduction
- 6. Structures sociales
- 7. Dynamique des groupes sociaux
- 8. Ontogenèse
- 9. Socialisation des jeunes
- 10. Modes de communication
- 11. Processus cognitifs
- 12. La culture chez les primates non humains
- 13. Protection et conservation des Primates
- 14. Bibliographie
PRIMATES
Article modifié le
Processus cognitifs
Depuis plusieurs années, la perception et plus particulièrement le traitement cognitif des signaux et des autres types d'information sont l'objet d'une attention particulière. L'analyse de la perception visuelle a été et est encore largement consacrée aux problèmes de discrimination et de catégorisation. C'est encore l'existence de réseaux relationnels durables, l'inertie de certains phénomènes sociaux tels que les relations de « dominance », et aussi leur remise en cause, qui démontrent l'importance de la perception visuelle au cours des interactions à courte portée. Elle contribue, pour un sujet, à la compréhension de la gestualité des partenaires, et, au-delà, à celle de l'ensemble de leurs relations. La description encore anecdotique de l'existence de « duperies » (deceipt des Anglo-Saxons), au cours d'interactions, renforce la conviction de l'existence de processus cognitifs complexes dans le traitement de l'ensemble des informations (Woodruff & Premack, 1979 ; Byrne & Whitten, 1987 ; De Waal, 1992).
C'est cependant par le biais de la communication sonore que se sont développées, au cours de ces dernières années, les études cognitivistes, grâce à une approche psycholinguistique du traitement du signal sonore. L'intérêt de ces études est qu'elles se déroulent à la fois au laboratoire et dans le milieu naturel des singes. La fonction de régulation de l'espacement de vocalisations fortes des cercocèbes (Lophocebus), des hurleurs et des gibbons a pu être démontrée par des expériences de « repasse » (playback) en milieu naturel (Waser, 1977 ; Whitehead, 1987 ; Mitani, 1985, respectivement). Mais cette technique a été aussi utilisée pour rechercher si une « perception catégorielle » existait chez les Primates et si les vocalisations, ou du moins certaines d'entre elles, pouvaient prendre valeur de symbole en « représentant » pour le récepteur un « référent » absent (non perceptible et/ou non perçu). Ces deux axes de recherches constituent, en fait, une approche comparative de la communication sonore des Primates et du langage humain.
La perception catégorielle
Les répertoires sonores de nombreuses espèces présentent un caractère gradué, c'est-à-dire qu'il existe, en particulier lors de séquences sonores, des gradations structurales réversibles ou non entre plusieurs classes vocales (Green, 1975 ; Deputte & Goustard, 1978 ; Robinson, 1979 b, Zimmerman, 1985). En dehors de ces séquences, toutes les classes vocales sont aisément différenciables. Il se pose alors le problème de savoir si et comment, lors de ces séquences, les gradations sont perçues, en d'autres termes si des catégories sont créées ou reconnues, ou si la séquence est perçue comme un seul, et nouveau, signal. Confrontés à un problème similaire, les Primates non humains procèdent-ils comme l'homme qui, écoutant le « continuum sonore » que constitue le langage parlé, le perçoit en le découpant de manière « catégorielle » ? Green (1975), chez le macaque japonais, a montré qu'il existait une relation claire entre les variations, graduées, subtiles, mais quantifiables au sein d'une même classe vocale (cris graves, tonals – côô sounds) et les contextes dans lesquels l'émetteur les produisait. Cela confère un caractère « phonétique » à ces cris (Snowdon, 1982). De son côté, Petersen (1982) a montré, chez cette même espèce, et en reprenant ces mêmes cris, que ces singes présentaient un avantage de l'oreille droite, donc de l'hémisphère gauche, dans le traitement des informations particulières et spécifiques contenues dans la modulation de ces côô sounds. Chez l'homme, ce même avantage existe lorsqu'un sujet traite les informations « linguistiques », alors que les informations paralinguistiques (hauteur des sons, intensité) sont traitées préférentiellement par l'hémisphère droit comme le sont les indices de reconnaissance visuelle (avantage de l'oreille gauche et du champ visuel gauche). Chez le macaque japonais, la perception du critère « hauteur du son » ne provoque ni attention spécifique ni latéralisation. Chez cette même espèce, Masataka (1983) montre que, pour des vocalisations structuralement voisines (cris d'alarme, vocalisations d'œstrus), c'est la hauteur de la fréquence terminale du cri qui est perçue de manière catégorielle : il existe deux types de réponse complètement opposées en deçà et au-delà du seuil de 8 kHz. Chez le marmoset pygmée (Cebuella pygmea), c'est la durée des cris de contacts (« trilles ») qui est perçue de manière catégorielle (Snowdon & Pola, 1978 ; Snowdon, 1982) : les sujets répondent aux trilles d'une durée inférieure à 250 millisecondes mais ne répondent plus au-delà de ce seuil précis.
La communication vocale symbolique
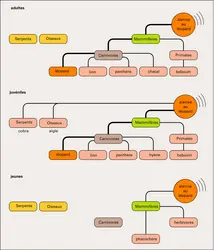
Catégorisation des prédateurs chez le vervet
Encyclopædia Universalis France

À cette recherche « phonétique » de la nature des vocalisations des Primates est associée une recherche « sémantique » : les cris des Primates constituent-ils, pour un récepteur spécifique, la représentation d'un stimulus, référent externe, même en son absence ? Dès 1967, Struhsaker, chez les vervets, Cercopithèques de savane, a montré que leurs cris d'alarme aux prédateurs variaient selon qu'ils avaient perçu un léopard, un serpent, un aigle ou un babouin. En reprenant cette analyse, Seyfarth et Cheney (1980) ont montré comment cette catégorisation s'établit au cours de l'ontogenèse et comment des types d'alarmes acquièrent une valeur sémantique : les enfants vervets émettent des alarmes « à l'aigle » pour tout ce qui vole, mais pas pour ce qui marche ; les juvéniles (entre un an et la maturité sexuelle) restreignent l'émission de ce type d'alarme, presque exclusivement, aux seuls rapaces, et enfin les adultes ne l'expriment qu'à la vue de l'aigle martial, le plus redoutable de leurs prédateurs. Une discrimination au niveau de l'espèce succède à des différenciations préalables en « classes ». De plus, les sujets répondent de manière adaptée à la perception de ces seuls cris d'alarme. Seyfarth et al. (1980) émettent alors l'hypothèse que ces cris, structuralement « discrets », c'est-à-dire très différents par leurs paramètres acoustiques, désigneraient les différentes classes de prédateurs prenant alors valeur de véritables symboles. Ces expérimentateurs diffusent ces cris en l'absence réelle de prédateurs, à des vervets qui se trouvent soit au sol, soit dans les arbres. Ils observent que, percevant l'alarme au léopard, les vervets courent dans les arbres ou y montent plus haut, et que, percevant l'alarme à l'aigle, ils fuient sous un couvert, sortent de l'arbre et regardent en l'air, enfin, à l'écoute de l'alarme au serpent, ils regardent au sol ou vers le bas. Ainsi le cri seul est susceptible de provoquer les mêmes réponses que lorsque le prédateur est réellement présent. Il se peut que les enfants vervets apprennent ces réponses adaptées en associant le cri perçu et la réponse qu'ils observent chez leur mère (ils peuvent même alors la vivre lorsqu'ils sont transportés !) ou chez d'autres partenaires. D'autres types de cris présentent des variations qui, bien qu'apparaissant dans un continuum, sont catégorisables : c'est le cas des cris aigus des macaques rhésus (Gouzoules et al., 1984), des grognements des vervets (Cheney & Seyfarth, 1982) ou des trilles des atèles (Masataka, 1986). Dans leur contexte habituel, les cris aigus des macaques sont généralement émis par des jeunes agressés par un congénère. Les réponses produites lors de la « repasse » de ces cris montrent que ces cris peuvent désigner aux membres du groupe le type de l'agresseur et la sévérité de l'attaque. De plus, la perception de caractéristiques individuelles conduit des femelles adultes à s'orienter directement vers la mère du jeune dont le cri a été diffusé, révélant ainsi la connaissance d'une relation. Chez beaucoup d'espèces de Primates, de nombreuses séquences vocales, y compris les duos, paraissent organisées syntactiquement, c'est-à-dire que la succession des cris qui les composent n'est pas aléatoire et paraît répondre à des « règles », mais rien ne permet d'affirmer que ces règles soient de même nature que celles qui gouvernent le langage humain (Robinson, 1984).
Les études constituant des approches cognitivistes du comportement des Primates continuent à se multiplier, qu'elles concernent la connaissance de leur environnement, non social ou social, notamment l'individualisation du partenaire (visuelle, vocale, olfactive). Plusieurs travaux ont démontré expérimentalement que les primates supérieurs (macaques, mangabeys) sont capables de reconnaître individuellement des partenaires qui leur sont présentés sur des écrans d'ordinateur (Dasser, 1987 ; Gomes da Silva & Deputte, 1998). La démonstration de cette reconnaissance impliquait que les sujets étaient capables de grouper, dans une même catégorie, différentes représentations du visage d'un même individu (face, profils, trois-quarts) et dans des catégories différentes ces représentations et celles d'autres individus. D'autres études ont abordé un phénomène plus complexe de reconnaissance, la reconnaissance multimodale : certains objets, mais tous les partenaires sociaux, sont susceptibles d'être reconnus selon plusieurs modalités sensorielles, notamment visuelle et vocale. La question est de savoir si les animaux testés sont capables d'associer un visage (image présentée sur un écran d'ordinateur) et une vocalisation (diffusée par un haut-parleur) appartenant à l'individu dont le visage est présenté, et cela quelle que soit la diversité des orientations des visages et de la nature des vocalisations diffusées. Certains éléments permettent de penser que cette reconnaissance bimodale, visuelle et acoustique, existe au moins chez les espèces qui ont été testées, c'est-à-dire les chimpanzés (Parr, 2004) et les mangabeys (Bovet et Deputte, 2006).
Le développement des études sur la cognition des primates s'inscrit dans le cadre défini par Griffin (1976) et déjà suggéré par Hinde (1966), celui d'accorder a priori des états mentaux aux espèces animales autres que Homo sapiens. Cette approche de Griffin envisage une continuité darwinienne aussi bien en ce qui concerne l'anatomie, la physiologie, mais aussi les phénomènes plus complexes du traitement des informations sociales et non sociales. Les espèces animales, notamment les primates, auraient toutes des « représentations » (Vauclair, 1987), des différences pouvant alors exister dans leur manière d'organiser, de rappeler et de communiquer ces représentations. La place de l'espèce Homo sapiens parmi les autres espèces de primates se trouve au centre des préoccupations scientifiques. Frans de Waal, en adoptant l'attitude de Griffin, a entrepris des études comparatives du comportement social de plusieurs espèces de macaques et des chimpanzés communs (De Waal, 1992). Il a analysé le comportement interactif de partenaires cinq minutes après la résolution d'un conflit qui avait éclaté entre eux. Il a montré qu'entre ces protagonistes les interactions qui suivaient immédiatement un conflit étaient qualitativement et/ou quantitativement différentes des interactions contrôles (interactions entre ces deux mêmes partenaires le lendemain, à la même heure que celle du conflit) et qu'elles étaient de nature amicale. Frans de Waal a alors défini ce phénomène comme étant de la « réconciliation », reprenant en cela, à dessein, un terme jusqu'alors utilisé uniquement chez Homo sapiens. Ce phénomène a pu être démontré chez plusieurs espèces de macaques, chez des vervets et chez les chimpanzés communs. Cette large distribution phylogénétique du phénomène laisse à penser que ce processus aurait été sélectionné dans certaines organisations sociales, pour « tamponner » l'effet disperseur d'une agression et ainsi maintenir la permanence de la cohésion des groupes, essence de la socialité. Par la formulation de ce phénomène de « réconciliation », chez les primates non humains, De Waal, a clairement souligné le caractère dual de toute relation sociale. Toute relation sociale présente deux composantes : l'une amicale et favorisant la cohésion ; l'autre agonistique, induisant une distanciation des partenaires. De nombreuses approches abordent le problème de la « théorie de l'esprit » (Theory of Mind, Premack et Woodruff, 1978) dans une perspective comparative. Les primates non humains, et particulièrement les chimpanzés communs mais aussi des capucins et des mangabeys, sont-ils capables d'attribuer des savoirs, voire des intentions à des congénères (Povinelli et al. 1990 ; Tomasello et al., 1999) ? Seuls les chimpanzés montrent cette « théorie de l'esprit » à un degré supérieur à celui des autres espèces de primates non humains (Povinelli et al., 1991), mais ce degré reste simple par rapport aux capacités d'un enfant de deux ans. Néanmoins, quel que soit le degré de ces capacités, les chimpanzés, mais aussi de nombreuses espèces de primates, sont susceptibles de développer, en particulier dans des contextes sociaux, des stratégies adaptatives très complexes (De Waal, 1982).
Accédez à l'intégralité de nos articles
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Bertrand L. DEPUTTE : docteur en éthologie, docteur ès sciences, professeur à l'École nationale vétérinaire d'Alfort
Classification
Médias

Singes douroucoulis
Gérard Lacz /Biosphoto/ Photononstop

Tamarin
Steve Clancy Photography/ Moment/ Getty Images

Ouistiti
Stuart Westmorland/ The Image Bank/ Getty Images
Autres références
-
BABOUIN
- Écrit par Marie-Claude BOMSEL
- 414 mots
- 2 médias

Singe au corps puissant, caractérisé par un museau allongé et nu comme celui du chien, d'où leur autre nom commun de cynocéphale (signifiant « tête de chien »). Répartition géographique : Afrique centrale, Afrique du Sud et Arabie. Habitat : savanes, collines rocheuses et forêts. Classe : Mammifères...
-
CERCOPITHÈQUE
- Écrit par Marie-Claude BOMSEL
- 436 mots
Singe de taille moyenne, au corps svelte et à longue queue non préhensile, vivant dans les forêts africaines, au sud du Sahara. Classe : Mammifères ; ordre : Primates ; famille : Cercopithécidés.
Autrefois appelés « guenons », les cercopithèques, représentés par plus de vingt espèces,...
-
CHASSEURS-CUEILLEURS (archéologie)
- Écrit par Jean-Paul DEMOULE
- 4 728 mots
- 3 médias

-
CHIMPANZÉ
- Écrit par Marie-Claude BOMSEL
- 639 mots
- 1 média

- Afficher les 28 références
Voir aussi
- MIMIQUE
- FÉCONDITÉ
- MONOGAMIE ANIMALE
- DÉVELOPPEMENT ANIMAL ou ONTOGENÈSE ANIMALE
- ESPRIT THÉORIE DE L'
- GLANDES
- PLACENTAIRES ou EUTHÉRIENS
- NEUROCRÂNE
- CYCLE ŒSTRAL
- SEXUEL COMPORTEMENT
- PLANTIGRADES
- PROSIMIENS
- LEMURIDAE ou LÉMURIDÉS
- INDRIDAE ou INDRIIDAE
- DAUBENTONIIDAE
- LORISIDÉS
- TARSIERS
- SINGES
- PLATYRHINIENS ou PLATYRRHINII
- CATARHINIENS ou CATARRHINII
- ANIMAL LANGAGE
- RÉPARTITION DES FLORES & DES FAUNES
- SIMIENS ou ANTHROPOIDEA
- ŒSTRUS
- EXTINCTION ou DISPARITION DES ESPÈCES
- CHEIROGALEIDAE
- GALAGIDAE
- ATÉLINÉS
- COLOBINÉS
- GROUPEMENTS ANIMAUX
- ANIMAL RÈGNE
- ANATOMIE ANIMALE
- POIL
- ONGLE
- PONGIDÉS ou PONGIDAE
- ODORANTES GLANDES
- ACCOUPLEMENT
- ANTHROPOMORPHES
- HYLOBATIDÉS ou HYLOBATIDAE
- ENVIRONNEMENT, droit et politique
- ATTACHEMENT THÉORIE DE L'
- ANTHROPISATION
- MÈRE-ENFANT RELATION
- POPULATIONS ANIMALES & VÉGÉTALES
- GESTATION
- HERBIVORES
- DOIGTS, zoologie
- CHANT, éthologie
- COMMUNICATION ANIMALE
- BIODÉMOGRAPHIE
- STÉRÉOTYPES COMPORTEMENTAUX
- MARQUAGE DU TERRITOIRE, éthologie
- ALARME, éthologie
- ATTRACTION, éthologie
- LÉMURIFORMES
- STREPSIRHINES
- HAPLORHINES
- CERCOPITHÉCOÏDES
- CERCOPITHÉCINÉS ou CERCOPITHÉCIDÉS
- GELADAS
- HOMINOÏDES
- PITHÉCINÉS
- RHINARIUM
- ATÉLIDÉS
- LORISIFORMES
- MEGALAPIDAE
- AYE-AYE
- ATÈLES
- CALLIMICO
- CALLITHRICIDÉS
- CÉBIDÉS
- MURIQUIS
- SAQUIS
- OUAKARIS
- MANGABEY
- NASIQUE
- PROTECTION ou CONSERVATION DES ESPÈCES
- ANIMAUX SAUVAGES EXPLOITATION DES
