
- 1. Le transfert d'information génétique au sein de la cellule
- 2. Rôle des facteurs de transcription
- 3. Autres niveaux de régulation de l'expression des gènes
- 4. Exemple de différenciation contrôlée par le contexte extracellulaire
- 5. Le programme de développement d'un organisme entier
- 6. Les modifications du programme
- 7. L'ambiguïté de la notion de programme
- 8. La reprogrammation
- 9. Bibliographie
PROGRAMME DE DÉVELOPPEMENT D'UN ORGANISME
L'ambiguïté de la notion de programme
La notion de programme génétique a été proposée dans les années 1970 afin de conceptualiser ce que l'on commençait tout juste à comprendre des mécanismes moléculaires régissant la différenciation cellulaire et le développement embryonnaire. Elle a émergé bien sûr des grandes découvertes de la biologie moléculaire, de la biochimie et de la génétique, mais aussi des concepts et techniques utilisés en informatique. Cette notion était tellement séduisante qu'elle s'est imposée, peut-être dangereusement, comme un dogme central.
En réalité, la comparaison entre un organisme vivant et un ordinateur tourne court assez rapidement. Seule la transcription d'un gène pris isolément correspond formellement à la lecture d'un programme en informatique. D'ailleurs, les chercheurs utilisent très fréquemment cette possibilité de pouvoir faire transcrire et traduire un gène eucaryote en dehors du contexte cellulaire, en l'introduisant soit dans un système artificiel totalement reconstitué, soit dans une bactérie. Les choses se compliquent nettement lorsqu'on se place dans le contexte tissulaire normal. En effet, le décodage du message génétique est effectué par un ensemble de protéines qui sont elles-mêmes synthétisées uniquement si une partie du message génétique a été lue ! De plus, ces protéines changent quantitativement et qualitativement au cours du développement embryonnaire, donc au fur et à mesure que le vaste programme censé diriger le développement est lu. Par conséquent, on se retrouve dans une situation où l'ordinateur permettant la lecture du programme évoluerait au fur et à mesure de cette lecture. En outre, plusieurs exemples nous ont montré que la lecture du programme dans une cellule peut permettre à celle-ci de modifier l'environnement de ses voisines, lesquelles verront alors en réponse leur programme se modifier. C'est probablement ce type de mécanisme qui permet un développement coordonné et synchronisé à l'échelle de l'organisme entier.
Enfin, il faut savoir que le programme génétique est capable de se diversifier au cours de la vie d'un individu, soit au hasard, soit pour remplir une fonction particulière. Les modifications au hasard peuvent survenir aussi bien dans les cellules somatiques que dans les cellules germinales et sont alors transmises à la descendance. L'environnement (rayons U.V., substances chimiques) ainsi que le mécanisme de réplication de l'ADN sont sources de nombreuses mutations au hasard. Cependant, celles-ci interviennent à très basse fréquence car elles sont généralement réparées par un ensemble d'enzymes constituant ce que l'on appelle le système SOS. D'autres modifications au hasard interviennent à fréquence plus élevée. Elles sont dues au déplacement de petites séquences mobiles, les transposons et les rétrotransposons, qui sont capables de s'exciser du génome et d'aller se réinsérer à peu près n'importe où. Leur transposition est source d'évolution : s'ils s'insèrent dans un gène, celui-ci deviendra probablement non fonctionnel. Au contraire, s'ils s'insèrent dans un promoteur ou un enhancer, on peut imaginer qu'ils modifient le niveau d'expression du gène. Plusieurs cas de maladies génétiques s'expliquent par des insertions de séquences mobiles. Par exemple, l'hémophilie A est due à l'insertion d'un rétrotransposon dans le gène du facteur VIII, un facteur de coagulation. De même, une des causes de la myopathie de Duchenne est l'inactivation, après insertion, du gène de la dystrophine. Les transposons et rétrotransposons sont des séquences fortement répétées. De ce fait, ils permettent des événements de recombinaison entre chromosomes non homologues. Ainsi s'explique la stérilité mâle XX, dans laquelle une partie du chromosome Y a été remplacée par recombinaison par un fragment de chromosome X. Le séquençage de l'ADN[...]
La suite de cet article est accessible aux abonnés
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Corinne ABBADIE : docteur en biologie du développement de l'université de Paris-VI, maître de conférences à l'université Lille I
Classification
Média
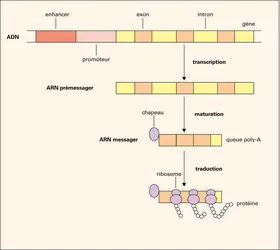
Transfert de l'information génétique
Encyclopædia Universalis France
