THERMORÉGULATION, biologie
Article modifié le
La température corporelle des homéothermes et sa régulation
La température corporelle n'est pas uniforme
La chaleur produite par le métabolisme cellulaire doit être transportée à la surface du corps pour pouvoir se dissiper vers l'extérieur, qui est en général à une température inférieure à celle du corps. La température de la surface du corps est donc inférieure à celle du centre, et cela permet le transfert de chaleur. En d'autres termes, la température du corps n'est pas uniforme. La production de chaleur par les tissus n'est pas davantage homogène, car certains organes (cœur, cerveau, viscères) en produisent bien plus que les autres, en particulier chez l'individu au repos. Nous pouvons donc, en première approximation, considérer le corps comme étant formé d'un noyau, dans lequel la plus grande partie de la production de chaleur prend place, et d'une enveloppe, qui comprend la peau et les muscles, et qui produit seulement une faible partie de la chaleur. Cela ne vaut que pour l'animal au repos. Lors d'un exercice musculaire, la situation est très différente, car le métabolisme augmente par un facteur 10, voire davantage. Cette augmentation correspond au fonctionnement des muscles (y compris les muscles respiratoires).
La température centrale de l'homme et des autres homéothermes montre des fluctuations journalières régulières de 1 à 2 0C. Les animaux diurnes montrent un maximum le jour et un minimum la nuit, les animaux nocturnes montrent des variations inverses. Ces cycles journaliers ont une origine endogène, et ils persistent pendant plusieurs semaines, même si l'animal est maintenu en éclairement continu. La température corporelle moyenne diffère entre les groupes de mammifères et d'oiseaux. Elle est, en particulier, nettement plus basse chez les monotrèmes et les autres groupes d'homéothermes « primitifs ».
La balance thermique des homéothermes

Thermogenèse des homéothermes
Encyclopædia Universalis France

Le métabolisme énergétique d'un homéotherme varie avec la température ambiante : il reste constant dans une zone plus ou moins large (zone de neutralité thermique) et devient plus élevé en deçà de cette zone (mise en jeu d'une thermogenèse régulatrice) et au-delà de cette zone (mise en jeu d'une thermolyse accrue).
La régulation thermique au froid
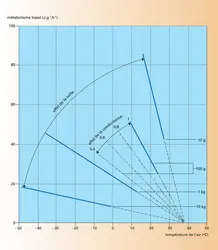
Métabolisme des mammifères lors de températures élevées
Encyclopædia Universalis France
Pour maintenir une température constante, un animal doit maintenir un état stationnaire, où la production de chaleur métabolique est égale aux pertes par conduction, convection, rayonnement et évaporation. Pour les températures relativement basses, l'évaporation correspond presque exclusivement aux pertes pulmonaires, qui ne représentent qu'un faible pourcentage du total. Il est possible de les négliger et d'écrire que Htot = Q = c ( (TC − TA), où Htot représente la chaleur produite par la thermogenèse de l'animal au repos (métabolisme basal), Q la chaleur perdue (thermolyse) et c un coefficient de conductance. Quand un animal change de milieu pour un milieu de température différente, il peut jouer sur l'un ou plusieurs des trois termes TC, c et Htot. Si nous ne considérons que le cas d'homéothermes, TC est constante (sauf en cas d'hibernation). Il ne reste donc que deux termes sur lesquels jouer, la production de chaleur Htot et la conductance c.
Augmentation de la production de chaleur (Htot)
L'augmentation de la production de chaleur peut prendre trois voies : des exercices musculaires volontaires, des tremblements involontaires des muscles (frissons thermiques) et une thermogenèse dans des tissus non musculaires. Les muscles ayant un rendement faible, une grande partie (> 80 p. 100) de l'énergie chimique qu'ils consomment est transformée en chaleur. Le travail musculaire produit donc une grande quantité de chaleur. Les frissons thermiques correspondent à des contractions simultanées de muscles antagonistes, qui se traduisent par des mouvements de faible amplitude. C'est un phénomène que chacun a pu expérimenter, de même que le recours à des exercices musculaires volontaires. Le cas le mieux connu de thermogenèse non musculaire correspond à celle qui s'effectue dans le tissu adipeux brun. Celui-ci est particulièrement important chez le nourrisson, qui ne possède pas encore les mécanismes du frisson thermique. Le tissu adipeux brun est également abondant chez les espèces qui hibernent, comme la marmotte, mais, en revanche, il est très réduit chez les adultes des espèces non hibernantes, l'homme compris. Les cellules du tissu adipeux brun sont riches en globules lipidiques, mais, à la différence du tissu adipeux normal, leur cytoplasme est rempli de mitochondries (c'est la forte concentration en cytochrome-oxydase qui donne sa couleur à ce tissu). Le tissu adipeux brun est très richement vascularisé. Le mécanisme de la thermogenèse met en jeu l'activation d'une lipase par la noradrénaline (récepteur β, agissant via une augmentation de l'AMP cyclique). Les acides gras formés sont dégradés par la β-oxydation et le cycle de Krebs, mais une protéine découplante de 32 kDa, la thermogénine, empêche la formation d'ATP par phosphorylation oxydative, ce qui fait que l'énergie produite est intégralement transformée en chaleur. La capacité de thermogenèse du tissu adipeux brun est supérieure à celle de tous les autres tissus, y compris le cœur.
Variations de la conductance (c) selon les espèces
Chez les mammifères, la fourrure représente une barrière pour les transferts de chaleur. Selon les espèces, son pouvoir isolant varie beaucoup. La fourrure des petits animaux est la moins épaisse, donc la moins isolante. Par ailleurs, ce sont ces mêmes animaux qui ont le rapport surface/volume le plus grand : les petits animaux compensent leur faible isolation en vivant dans un terrier où le microclimat est plus clément et/ou en hibernant. L'épaisseur de la fourrure varie avec les saisons (pelages d'été et d'hiver). Cela est surtout vrai pour les gros animaux, dont la conductance peut varier d'un facteur 2 selon les saisons. Chez les oiseaux, il existe des phénomènes comparables pour le plumage. La fourrure n'est bien isolante que sèche : humide, en particulier dans l'eau, elle perd une grande partie de son pouvoir protecteur, tandis que le milieu ambiant, plus conducteur, accélère encore le refroidissement. Dans ce cas, le facteur isolant principal n'est plus la fourrure, mais la couche adipeuse sous-cutanée. Les oiseaux aquatiques enduisent leurs plumes de substances hydrophobes pour les rendre non mouillantes et pour conserver ainsi au plumage son pouvoir isolant, même dans l'eau.
Le comportement des animaux est également important pour limiter leur refroidissement : les animaux replient les pattes sous le corps pour réduire la surface exposée. Ils peuvent aussi se regrouper les uns contre les autres, ce qui réduit la surface exposée par chacun. Ils peuvent enfin laisser la couche externe (l'enveloppe) du corps se refroidir (grâce à des mécanismes de vasoconstriction périphérique), ce qui diminue le gradient, donc le flux de chaleur.
Cas des mammifères aquatiques (notion d'hétérothermie régionale)
Les phoques et les baleines vivent dans des eaux froides. La conduction thermique de l'eau est, on l'a vu, environ vingt-cinq fois plus élevée que celle de l'air, et la convection liée au mouvement des animaux augmente encore son pouvoir refroidissant. Ces animaux possèdent une isolation thermique renforcée, réduisant leurs pertes de chaleur grâce à la présence d'une épaisse couche de lard sous-cutané, qui représente une fraction très importante du volume de l'animal. Dans l'eau, la température cutanée de l'animal est égale à celle de l'eau, et la température croît progressivement dans l'épaisseur de la couche de lard. Lorsque le phoque est à terre, sa fourrure l'isole bien, et sa température cutanée est bien plus forte. Fourrure et lard sont donc fonctionnellement très différents, en raison des possibilités qui existent de court-circuiter l'isolation par le lard.
Dans les membres, qui sont moins bien isolés que le reste du corps, chez le dauphin par exemple, il existe un système à contre-courant (chaque artère est entourée par des veines où le sang circule en sens inverse). Ce système crée un gradient thermique le long du membre, ce qui permet de réduire fortement les pertes de chaleur par les extrémités. De tels gradients thermiques existent également le long des membres chez les mammifères terrestres et des oiseaux, ainsi qu'au niveau des fosses nasales. On pourra noter l'existence, dans ce cas, d'une adaptation des fibres nerveuses sur leur trajet : ce ne sont pas les mêmes enzymes qui sont présentes le long des fibres, les extrémités contenant des formes moléculaires actives à basse température.
La régulation thermique au chaud
Nous avons vu que les homéothermes augmentent leur production de chaleur au-dessous d'une température critique. Au-dessus de cette valeur, le métabolisme reste constant dans la zone dite de thermoneutralité, zone dans laquelle de petites modifications de conductance permettent un équilibre, de façon que la chaleur soit dissipée aussi vite qu'elle est produite. Lorsque la température ambiante augmente, la différence (TC − TA) diminue, ce qui fait que les pertes de chaleur par conduction-convection et par rayonnement deviennent de plus en plus limitées. Dans l'égalité Htot = HC + HR + HS + HE, comme Htot reste constante (métabolisme basal) et que (HC + HR) diminue, cela signifie que (HS + HE) augmente. Si la température reste constante (HS = 0), on voit que HE augmente, donc l'animal fait de plus en plus appel aux mécanismes de thermolyse par évaporation. Lorsque TA = TC, Htot = HE : c'est alors uniquement l'évaporation qui permet le refroidissement de l'animal. C'est bien sûr encore le cas si TA > TC, auquel cas l'animal reçoit en plus de la chaleur de son environnement.
L'importance de la taille de l'animal
Un petit objet placé au soleil s'échauffe plus vite qu'un gros. Cela illustre l'importance du rapport S/V (surface/volume). Si l'on prend une sphère de rayon r, sa surface S est 4 πr2 et son volume est V = (4/3)π ( r3, donc son rapport S/V = 1/3r. La surface corporelle des mammifères est environ deux fois celle d'une sphère de même volume, donc S/V est aussi une fonction de 1/r. Il est donc clair que le rapport S/V des animaux diminue lorsque leur taille augmente. Dans le cas d'un animal placé dans un environnement chaud, le gain de chaleur par conduction et rayonnement est proportionnel à S (de même que Htot, car le métabolisme basal est lui aussi approximativement proportionnel à la surface du corps). La quantité de chaleur que l'animal doit perdre (par évaporation) est donc proportionnelle à S, ce qui place le petit animal dans des conditions très défavorables : si un homme placé dans le désert doit évaporer 1 litre de sueur par heure (soit 0,6 l/m2), soit environ 1,5 p. 100 de l'eau que contient son organisme, un petit rongeur devrait évaporer de 15 à 30 p. 100 de son poids pendant le même laps de temps, ce qui n'est guère envisageable. Les petits animaux doivent donc éviter la chaleur, et, pour cela, ils se réfugient dans leur terrier durant la plus grande partie du jour. Ceux qui sont actifs de jour, comme le spermophile, ne font que de courtes sorties au cours desquelles leur température corporelle atteint 42 0C, et ils retournent régulièrement dans leur terrier pour se refroidir. Les grands animaux comme le chameau et le dromadaire tirent un avantage du simple fait de leur grande taille (faible rapport S/V). Par ailleurs, ils peuvent laisser leur température descendre la nuit et augmenter le jour (HS), ce qui abaisse le gradient thermique et, surtout, permet de retarder la mise en œuvre des mécanismes de thermorégulation par évaporation.
Les modes d'évaporation
Placé dans un environnement chaud, un homme transpire. Cette sudation est le fait de glandes sudoripares réparties sur toute la surface du corps. Les chiens sont dépourvus de glandes sudoripares, et ils se refroidissent par évaporation au niveau de la partie supérieure du tractus respiratoire (par halètement ou polypnée thermique). Il existe une troisième méthode, qui est le léchage des membres. La sudation est courante chez les mammifères ; toutefois, la présence d'une fourrure ralentit les processus d'évaporation, ce qui explique que les animaux à pelage épais et vivant en milieu chaud aient plutôt recours à la polypnée. La polypnée existe chez des mammifères de grande taille, herbivores et carnivores ; elle existe également chez les oiseaux. Elle s'effectue à un rythme constant pour une espèce donnée, rythme qui correspond à la fréquence des oscillations élastiques de la cage thoracique, ce qui limite la dépense énergétique musculaire correspondante. La salivation et le léchage sont surtout pratiqués par les marsupiaux.
On pourra également noter que la sudation représente, en plus de la perte d'eau, une perte de sels, ce qui n'est pas le cas de la polypnée. La polypnée ne provoque pas d'hyperventilation (qui augmenterait la vitesse d'élimination du CO2 et provoquerait ainsi une alcalose respiratoire), car la fréquence élevée des mouvements respiratoires s'accompagne d'une diminution du volume d'air inspiré à chaque mouvement. Un autre intérêt de la polypnée est de permettre un refroidissement du sang qui irrigue le cerveau, grâce à la présence d'un échange thermique entre les veines du nez et les artères carotides. Ce dispositif existe également chez les reptiles, mais il est absent chez de nombreux mammifères, dont les rongeurs et les primates.
Les contrôles de l'homéothermie
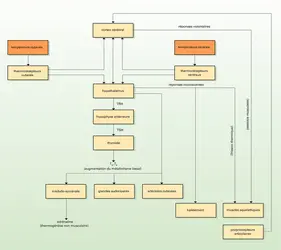
Homéothermes : mécanismes thermorégulateurs
Encyclopædia Universalis France
Nous venons de voir qu'il existe un grand nombre de mécanismes effecteurs : réponses vasomotrices, frisson thermique, thermogenèse du tissu adipeux brun, polypnée, sudation, etc. Ces différents mécanismes sont régulés de façon intégrée afin de maintenir constante la température du corps. Cela implique l'existence de récepteurs thermosensibles et de centres intégrateurs.
Il existe des récepteurs thermiques centraux situés dans l' hypothalamus (dans les aires préoptiques), qui joue en même temps le rôle de thermostat. Ce rôle a été établi par Henry Barbour dès 1912, grâce à des expériences d'implantation de thermodes chez un chien : la modification locale de la température provoquée par l'expérimentateur déclenchait une réponse thermorégulatrice appropriée. On en conclut que c'est donc la température du sang qui irrigue l'hypothalamus qui provoque la mise en jeu des mécanismes régulateurs. Ceux-ci empruntent pour l'essentiel des voies nerveuses, qui agissent, par exemple, sur la contraction des muscles squelettiques (frissons thermiques), le diamètre des artérioles cutanées ou la sécrétion des glandes sudoripares.
L'activité du thermostat central est modulée sous l'action d'afférences nerveuses en provenance de thermorécepteurs périphériques, en particulier ceux qui sont présents sur la peau (et particulièrement abondants sur la langue, le scrotum et les mamelles). Ces thermorécepteurs correspondent à des terminaisons axonales sensibles au chaud ou au froid, dont l'activité électrique varie avec la température. Leur excitation est transmise par des voies ascendantes vers l'hypothalamus, et elle est susceptible de déclencher une réponse thermorégulatrice, alors même que la température centrale n'a pas été modifiée. De même, la mise en jeu de propriocepteurs lors des mouvements musculaires déclenche le mécanisme de halètement dès le début de l'exercice chez l'animal.
À côté des voies nerveuses, il existe également des régulations hormonales. Celles-ci ont une importance souvent secondaire. On notera le rôle des hormones thyroïdiennes, qui stimulent la thermogenèse en agissant sur l'activité métabolique de l'ensemble des tissus, et celui de l' adrénaline produite par la médullo-surrénale, qui stimule la thermogenèse dans le tissu adipeux brun.
Les hormones peuvent aussi intervenir dans des mécanismes adaptatifs plus lents : c'est le cas des hormones thyroïdiennes, qui augmentent le métabolisme basal et donc la production de chaleur, et dont la concentration est accrue lors de l'acclimatation au froid. Dans le même ordre d'idées, les minéralocorticoïdes (l'aldostérone) interviennent dans la réduction de la teneur de la sueur en sels lorsque les individus commencent à s'acclimater à une température élevée. Ainsi, la perte de sels en cas de sudation abondante est considérablement réduite.
La thermorégulation met également en œuvre une régulation par voie nerveuse du comportement des animaux, conduisant, chez l'animal, à une modification des surfaces exposées, et, chez l'homme, à toute une série de comportements plus élaborés relatifs, par exemple, au choix de ses vêtements ou à la climatisation de son habitat.
Accédez à l'intégralité de nos articles
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- René LAFONT : professeur des Universités
Classification
Médias
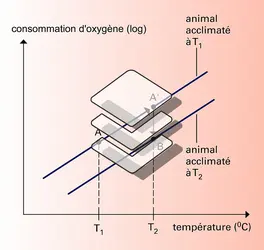
Température corporelle et variation du métabolisme
Encyclopædia Universalis France
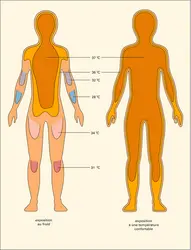
Isothermes de l'enveloppe de l'homme
Encyclopædia Universalis France
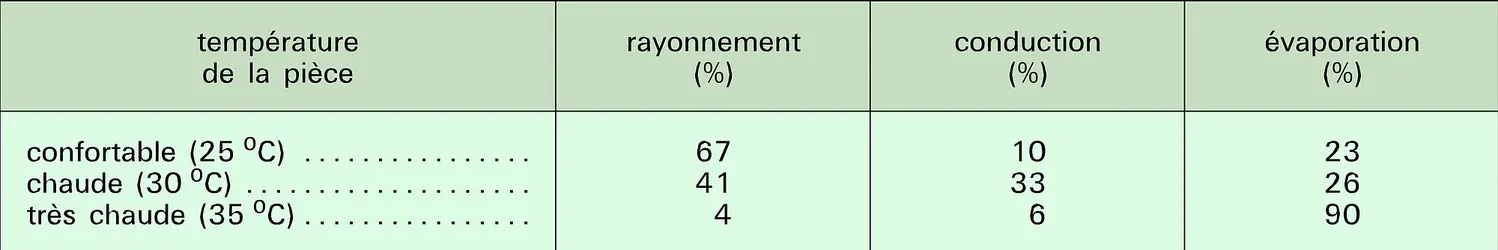
Pertes par rayonnement, conduction et évaporation
Encyclopædia Universalis France
Autres références
-
DÉSERTS
- Écrit par Roger COQUE , François DURAND-DASTÈS , Huguette GENEST et Francis PETTER
- 20 890 mots
- 16 médias
 ...sont pas compatibles avec la vie animale. Des animaux typiquement désertiques exposés en plein soleil aux heures chaudes de la journée meurent rapidement. Tous les animaux « à sang chaud » sont pourvus d'organes régulateurs qui leur permettent de maintenir leur température corporelle à un niveau normal voisin...
...sont pas compatibles avec la vie animale. Des animaux typiquement désertiques exposés en plein soleil aux heures chaudes de la journée meurent rapidement. Tous les animaux « à sang chaud » sont pourvus d'organes régulateurs qui leur permettent de maintenir leur température corporelle à un niveau normal voisin... -
DINOSAURES
- Écrit par Eric BUFFETAUT
- 7 345 mots
- 16 médias
 ...ont beaucoup varié au fil du temps. Les paléontologues les ont longtemps considérés comme de simples versions énormément agrandies de reptiles actuels. De ce fait, leur physiologie aurait été de type reptilien, c'est-à-dire que la température de leur corps variait en fonction de celle de leur milieu ;...
...ont beaucoup varié au fil du temps. Les paléontologues les ont longtemps considérés comme de simples versions énormément agrandies de reptiles actuels. De ce fait, leur physiologie aurait été de type reptilien, c'est-à-dire que la température de leur corps variait en fonction de celle de leur milieu ;... -
ÉCOLOGIE
- Écrit par Patrick BLANDIN , Denis COUVET , Maxime LAMOTTE et Cesare F. SACCHI
- 20 638 mots
- 15 médias
 ...statistiques, des relations entre certaines caractéristiques physiologiques ou morphologiques et certains facteurs du milieu, en particulier la température. Chez les homéothermes, par exemple, les tailles les plus grandes se rencontrent dans les régions les plus froides (règle de Bergmann). Cela est lié au...
...statistiques, des relations entre certaines caractéristiques physiologiques ou morphologiques et certains facteurs du milieu, en particulier la température. Chez les homéothermes, par exemple, les tailles les plus grandes se rencontrent dans les régions les plus froides (règle de Bergmann). Cela est lié au... -
ÉPIPHYSE ou GLANDE PINÉALE
- Écrit par Berthe VIVIEN-ROELS
- 2 325 mots
- 1 média
Des corrélations étroites ont également été établies chez de nombreuses espècesentre le développement de la glande pinéale et le degré d'homéothermie. De plus, il est démontré que la glande pinéale et les organes qui lui sont souvent associés chez les Vertébrés inférieurs (œil pariétal des lézards,... - Afficher les 18 références
Voir aussi
- CONGÉLATION
- CONVECTION ou CONVEXION
- POÏKILOTHERMIE ou PŒCILOTHERMIE
- ECTOTHERMIE
- ENDOTHERMIE
- BALANCE THERMIQUE, physiologie
- THERMOGENÈSE
- TRANSPIRATION ANIMALE ou SUDATION ANIMALE
- SUDORALES GLANDES
- THERMORÉCEPTEUR
- PHYSIOLOGIE
- ÉVAPORATION, physique
- TEMPÉRATURE
- POLYPNÉE
- HYPERTHERMIE
- ADIPEUX BRUN TISSU
- THERMOLYSE
- ACCLIMATEMENT
- BIOÉNERGÉTIQUE
- MUSCULAIRE TRAVAIL
- HOMÉOTHERMIE
- ADAPTATION BIOLOGIQUE
- AQUATIQUE VIE
- RAYONNEMENT THERMIQUE
- TAILLE CORPORELLE
- CONDUCTION THERMIQUE
