VISION Neurophysiologie visuelle
Article modifié le
Les structures sous-corticales
Projections rétiniennes
Les axones des cellules ganglionnaires convergent dans la papille (aussi appelée tache aveugle) pour former le nerf optique qui se projette sur plusieurs structures sous-corticales. La grande majorité (90 %) de ces axones se dirigent vers le corps genouillé latéral dorsal (CGL), un noyau (ensemble compact de neurones) qui fait partie du thalamus (ensemble de noyaux sous-corticaux fortement connectés au cortex de façon bidirectionnelle). Ils proviennent de neurones de type alpha, bêta et gamma qui se répartissent de façon différente dans les différentes couches cellulaires du CGL (cf. infra). Les autres cibles de la rétine sont : le colliculus supérieur, qui reçoit environ 10 % des fibres rétiniennes de type alpha et gamma, impliqué dans les mouvements des yeux et est la cible de fortes connexions du cortex visuel ; d'autres noyaux qui reçoivent des fibres de neurones gamma :
– le noyau suprachiasmatique, une structure de l'hypothalamus participant à la régulation des rythmes circadiens par la lumière ;

– certains noyaux du prétectum impliqués dans la motricité oculaire réflexe et l'accommodation ;
– les noyaux du système optique accessoire qui sont mis en jeu dans les réflexes de stabilisation du globe oculaire lors de mouvements de la tête.
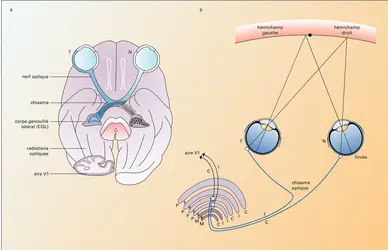
Voies optiques et projections rétiniennes au CGL
Encyclopædia Universalis France
Les fibres rétiniennes qui se dirigent vers le CGL subissent une décussation, c'est-à-dire un entrecroisement, partielle au niveau du chiasma optique qui a pour effet de restreindre la représentation de chaque hémichamp visuel à l'hémisphère cérébral opposé (controlatéral). En effet, le champ visuel peut être divisé en deux hémichamps par un plan vertical passant par le nez et le point de fixation des yeux. De la même façon, chaque rétine peut être divisée en deux hémirétines correspondantes. Un point de chaque hémichamp se projette dans l'hémirétine temporale de l'œil du côté controlatéral et dans l'hémirétine nasale de l'œil du même côté (ipsilatéral). Les fibres rétiniennes provenant des deux yeux sont réorganisées dans le chiasma optique de façon à ce que les fibres rétino-genouillées provenant de l'hémirétine nasale croisent pour innerver le CGL controlatéral, alors que celles qui proviennent de l'hémirétine temporale se projettent dans le CGL ipsilatéral.
Le corps genouillé latéral
La plupart des neurones de cette structure (80 à 90 %) reçoivent des afférents rétiniens et envoient un axone dans les radiations optiques vers le cortex de l'aire V1. Ce sont les neurones relais. Les autres neurones de cette structure sont des neurones inhibiteurs à projection locale. Un des rôles supposés du CGL est de permettre la séparation des neurones innervés par les cellules rétiniennes de type alpha, bêta et gamma en différentes couches cellulaires et de mettre en registre les représentations de l'hémichamp visuel controlatéral pour chacune de ces catégories. Ce rôle de tri et de mise en registre est en accord avec sa forme laminaire caractéristique du CGL.
Une section du corps genouillé latéral dorsal comporte six couches de neurones. Chaque couche contient une représentation de l'hémichamp visuel controlatéral par des neurones innervés par des axones provenant de cellules ganglionnaires de l'un ou l'autre des deux yeux. Certaines couches sont innervées par les cellules ganglionnaires de la partie nasale de l'œil controlatéral. Les autres couches reçoivent des afférents en provenance de l'hémirétine temporale de l'œil ipsilatéral. L'innervation du CGL est organisée de telle façon que les champs récepteurs des neurones situés dans des couches adjacentes recouvrent la même région du champ visuel. De ce fait, une petite tache de lumière située dans un hémichamp visuel active des neurones alignés le long d'une ligne perpendiculaire aux couches du CGL controlatéral. Les axones des connexions en retour (ou feed-back) du cortex visuel traversent les différentes couches du CGL selon ce même axe, permettant ainsi une modulation de l'activité de l'ensemble des neurones activés par ce stimulus.
Les neurones relais du corps genouillé latéral peuvent être divisés en trois catégories : les neurones P, qui appartiennent aux quatre couches supérieures appelées parvocellulaires ; les neurones M, qu'on trouve dans les deux couches inférieures dites magnocellulaires ; les neurones K (abréviation de koniocellulaires), qui sont situés entre les couches. Les neurones P, innervés par les cellules bêta de la rétine, relaient l'information concernant la vision des couleurs et des détails fins. Leurs champs récepteurs sont de petite taille et sont sensibles aussi bien aux contrastes de luminance qu'aux contrastes de couleur. Les neurones M sont innervés par les cellules alpha de la rétine. Ils possèdent une moins bonne acuité et ne codent pas la couleur car ils donnent la même réponse à des stimuli de couleurs différentes s'ils ont la même luminance. Contrairement aux neurones P, les neurones M répondent de façon vigoureuse à de hautes fréquences temporelles (images en mouvement rapide sur la rétine ou papillotement rapide de tache lumineuse) et à de faibles contrastes (le contraste est le rapport de l'augmentation de la luminance du stimulus par rapport à la luminance du fond). Enfin, les neurones K, qui sont situés entre les couches principales du CGL, relaient l'information provenant des neurones gamma qui se projettent dans le thalamus et ont des propriétés mal connues (certains sont sensibles au contraste jaune-bleu).
Une autre différence importante entre les neurones M, P et K concerne la latence de leur réponse aux stimuli visuels : les cellules M commencent à répondre en moyenne 50 millisecondes après la présentation d'un stimulus, les cellules P répondent 20 millisecondes après les M, et les neurones K sont retardés de 10 millisecondes par rapport aux cellules P. On ne note pas de transformation majeure des propriétés des neurones dans le CGL par rapport à leurs afférents rétiniens : par exemple, les neurones P de type on et off ont des propriétés similaires aux neurones correspondant dans la rétine (cellules de type bêta on et off).
La ségrégation des classes de neurones P, M et K dans différentes couches du CGL constitue un bon exemple d'un principe général d'organisation du système visuel, celui du regroupement de neurones ayant des propriétés fonctionnelles voisines. Pour des raisons de simplification de la connectivité, l'information provenant de la rétine est séparée au niveau du CGL en plusieurs groupes fonctionnels qui cheminent en parallèle et de façon relativement indépendante. On parle de canaux parallèles de transfert et de traitement. L'information codée par les différents canaux se recouvre largement : les neurones P, M et K sont activés ensemble par tous les stimuli visuels, sauf dans des cas exceptionnels comme les expériences de laboratoire. En effet, ce n'est que dans le cas de stimuli contenant de très hautes fréquences spatiales (au-delà de 30 cycles par degré d'angle visuel), ou de stimuli colorés de même luminance, que les neurones P sont les seuls à répondre. Ce n'est que pour des contrastes très faibles (au-dessous de 8 %) ou de hautes fréquences temporelles (au-dessus de 30 Hz) que le canal M est le seul à fonctionner.
Cette ségrégation fonctionnelle dans les différentes couches du CGL a permis de tester le rôle de ces diverses classes en réalisant chez le singe des expériences de lésion ou d'inactivation sélective : à la suite de lésions des couches parvocellulaires, on observe une perte de la vision des couleurs, de forts déficits dans la vision des formes, des détails et de la profondeur. Des lésions des couches magnocellulaires n'ont pas d'effets sur l'acuité, la perception des formes ou des couleurs mais abolissent la capacité du singe à percevoir une tache de lumière papillotant à haute fréquence et des mouvements rapides de stimuli (P. H. Schiller, N. K. Logothetis et al., 1990).
Outre les afférences rétiniennes, les neurones du CGL reçoivent également un petit contingent d'afférences en provenance de la formation réticulée mésencéphalique et de structures du tronc cérébral (raphé dorsalis et locus coeruleus). Ces afférences adaptent le niveau de transmission des messages rétiniens vers le cortex selon les différents états d'éveil. Cependant, le plus gros contingent de fibres afférentes au CGL est constitué par des voies en retour (feed-back) provenant de l'aire V1 (dix fois plus que d'afférents rétiniens). Le rôle de ces voies en retour demeure mal connu.
Il existe un autre noyau thalamique qui est impliqué dans la vision, le pulvinar. Il ne reçoit pas de connexions de la rétine mais est fortement connecté de façon réciproque avec les différentes aires corticales visuelles. Sa taille est particulièrement importante chez l'homme, mais son rôle dans la vision reste largement mystérieux.
Accédez à l'intégralité de nos articles
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Jean BULLIER : directeur de recherche au C.N.R.S., directeur du centre de recherche cerveau et cognition, université Paul-Sabatier, Toulouse
Classification
Médias
Voies optiques et projections rétiniennes au CGL
Encyclopædia Universalis France
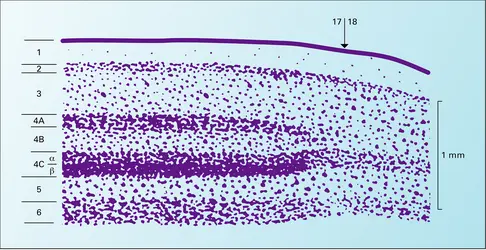
Cortex cérébral (aires 17-18)
Encyclopædia Universalis France

Vision humaine : îlots à cytochrome oxydase abondante dans l'aire V1, modules d'orientation et dominance oculaire
2002 by Society for Neuroscience
Autres références
-
AMPHIBIENS ou BATRACIENS
- Écrit par Pierre CLAIRAMBAULT , Philippe JANVIER et Jean-Claude RAGE
- 6 177 mots
- 19 médias
 ...suffit en effet de comparer une grenouille et un triton). Chez les Gymniophones, l'œil se trouve sous la peau ou même sous la couverture osseuse du crâne. Certains amphibiens, comme le protée ou les Gymniophones, sont aveugles (adaptation à la vie souterraine). La protection de l'œil est assurée par la présence...
...suffit en effet de comparer une grenouille et un triton). Chez les Gymniophones, l'œil se trouve sous la peau ou même sous la couverture osseuse du crâne. Certains amphibiens, comme le protée ou les Gymniophones, sont aveugles (adaptation à la vie souterraine). La protection de l'œil est assurée par la présence... -
CERVEAU ET GESTES
- Écrit par Didier LE GALL et François OSIURAK
- 916 mots
- 1 média
 Les informations visuelles permettent de déterminer la position d’un objet par rapport au corps. Les informations proprioceptives proviennent de capteurs situés au niveau des tendons, des articulations et des muscles. Grâce à ces informations, nous sommes capables à tout moment d’avoir une représentation...
Les informations visuelles permettent de déterminer la position d’un objet par rapport au corps. Les informations proprioceptives proviennent de capteurs situés au niveau des tendons, des articulations et des muscles. Grâce à ces informations, nous sommes capables à tout moment d’avoir une représentation... -
COGNITIVES SCIENCES
- Écrit par Daniel ANDLER
- 19 265 mots
- 4 médias
 Les recherches sur lavision sont peut-être la branche la plus « scientifique » (au sens étroit) des sciences cognitives. C'est aussi celle dans laquelle les neurosciences jouent le plus grand rôle. Le second fait n'explique qu'en partie le premier : la vision présente par rapport à d'autres modalités...
Les recherches sur lavision sont peut-être la branche la plus « scientifique » (au sens étroit) des sciences cognitives. C'est aussi celle dans laquelle les neurosciences jouent le plus grand rôle. Le second fait n'explique qu'en partie le premier : la vision présente par rapport à d'autres modalités... -
COULEUR
- Écrit par Pierre FLEURY et Christian IMBERT
- 7 733 mots
- 21 médias
 On considère en psychophysiologie, sous les noms de teinte, saturation et luminosité d'une lumière, trois qualités dont chacune dépend principalement de la caractéristique « chromatique » (λd ou p) ou photométrique (L) correspondante, mais parfois aussi quelque peu des deux autres....
On considère en psychophysiologie, sous les noms de teinte, saturation et luminosité d'une lumière, trois qualités dont chacune dépend principalement de la caractéristique « chromatique » (λd ou p) ou photométrique (L) correspondante, mais parfois aussi quelque peu des deux autres.... - Afficher les 43 références
Voir aussi
- FORMES RECONNAISSANCE DES
- RÉTINE
- POTENTIEL ÉLECTRIQUE
- LUMINANCE
- CELLULES GANGLIONNAIRES RÉTINIENNES
- CELLULES BIPOLAIRES RÉTINIENNES
- NERF OPTIQUE
- ÉCLAIREMENT
- CHIASMA OPTIQUE
- VISION NOCTURNE ou VISION SCOTOPIQUE
- PHOTORÉCEPTEUR
- PHYSIOLOGIE
- AXONE ou CYLINDRAXE
- POTENTIEL DE REPOS
- SENSORIELLES AIRES
- POTENTIEL D'ACTION
- EXCITABILITÉ
- CONTRASTE
- PAPILLE OPTIQUE
- NEUROPHYSIOLOGIE
- CHAMP RÉCEPTEUR, neurophysiologie
- CELLULES AMACRINES
- DOMINANCE OCULAIRE
- MOUVEMENT PERCEPTION DU
- SÉLECTIVITÉ À L'ORIENTATION, neurophysiologie
- CELLULES HORIZONTALES RÉTINIENNES
- NEURONE ou CELLULE NERVEUSE
- FOVEA
- ACUITÉ VISUELLE
- CORPS GENOUILLÉ
- CÔNE, cytologie
- BÂTONNET, cytologie
- INHIBITION
- RÉSEAU NERVEUX
- PROJECTIONS, neurophysiologie
- AFFÉRENCES, neurologie
- PERCEPTION VISUELLE
- CHAMP VISUEL
- HYPERPOLARISATION, neurobiologie
- VISION DES COULEURS
- LOBE PARIÉTAL
