
- 1. Notion d’activité cérébrale
- 2. Visualiser l’activité électrique des neurones
- 3. L’imagerie cérébrale par électroencéphalographie
- 4. L’imagerie cérébrale fonctionnelle par radioéléments
- 5. Imagerie par magnétométrie
- 6. Imagerie par résonance magnétique nucléaire
- 7. L’imagerie multiple par combinaison des techniques
- 8. Définition des activités cérébrales à échelle locale
- 9. Convergence et divergence des données d’imagerie cérébrale
- 10. Difficulté de la projection sur l’homme de résultats obtenus sur des modèles animaux
- 11. Construction théorique à partir des données d’imagerie
- 12. Nécessité des modèles animaux
- 13. L’imagerie pour la clinique neurologique
- 14. L’imagerie, l’éthologie et la psychothérapie
- 15. Bibliographie
VISUALISATION DE L'ACTIVITÉ DU CERVEAU
Convergence et divergence des données d’imagerie cérébrale
Lorsqu’il s’agit d’étudier à l’échelle des grandes aires corticales les localisations des régions impliquées dans certaines tâches cognitives, il arrive souvent que différentes techniques aboutissent à des résultats très similaires. Cela est le cas par exemple pour la caractérisation des aires cérébrales impliquées dans l’analyse syntactique (grammaticale) des phrases par la MEG et l’IRMf, en accord avec les études sur des patients présentant des lésions de ces régions et chez lesquels on peut mettre en évidence des troubles fonctionnels correspondants. Pour la fonction de reconnaissance des visages, l’étude des régions cérébrales impliquées bénéficie également des études d’IRMf, qui ont confirmé les résultats électrophysiologiques très nombreux obtenus chez l’animal sur la localisation des neurones impliqués dans cette fonction.
Cependant, ces accords ne s’établissent que sur des corrélations simples entre aires cérébrales et fonctions, et ne présagent en rien d’un quelconque consensus sur les mécanismes cérébraux impliquant ces aires qui rendraient compte de la fonction étudiée. Cela tient en partie au fait que ces correspondances ne sont établies que pour des intervalles de temps de quelques centaines de millisecondes qui ne permettent pas de corrélations dans les processus cognitifs rapides, qu’il faudrait néanmoins connaître pour comprendre réellement les mécanismes neuronaux en jeu, un peu comme la biochimie des années 1920 s’était attaquée à la chimie des réactions secondaires du métabolisme non plus simplement pour décrire certaines fonctions cellulaires, mais pour créer de véritables modèles biochimiques.
Par ailleurs, des désaccords sur l’activation ou pas de certaines sous-aires cérébrales lors d’une même tâche cognitive peuvent apparaître selon les techniques utilisées. Les raisons de telles divergences sont extrêmement diverses et nombreuses. D’une part, certaines techniques peuvent connaître des limitations opérationnelles lorsqu’il s’agit de visualiser certaines activités dans certaines régions anatomiques précises – ainsi l’IRMf, la MEG et l’EEG dans des conditions où les signaux sont masqués (souvent par annulation de signaux opposés dans une même région ou par des masquages dus à des processus physiologiques non neuronaux). D’autre part, les mesures d’imagerie étant réalisées de façon globale à l’intérieur d’un petit volume de tissu cérébral analysé, il est courant qu’une activité soit masquée en raison de la présence de plusieurs réseaux de neurones mêlés, alors que les enregistrements électriques de neurones individuels dans les mêmes régions détectent des activations de populations hétérogènes de neurones.
Enfin, la comparaison des résultats entre différentes techniques est conditionnée par leur alignement spatial dans un même repère tridimensionnel, facilité par l’implémentation de la technique de repérage spatial de la stéréotaxie pour chaque technique d’imagerie. L’alignement avec les données microscopiques de cytoarchitectonie est particulièrement difficile, mais de récentes études montrent que l’emploi de l’IRM de diffusion est parfois très efficace pour aligner les données d’imagerie fonctionnelle avec celles-ci, par alignement des faisceaux d’axones. Ces derniers peuvent en effet constituer localement des minirepères à l’échelle du dixième de millimètre, et donc à celle des petites populations neuronales des régions cérébrales bien différenciées anatomiquement et fonctionnellement à cette échelle.
La suite de cet article est accessible aux abonnés
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Jean-Gaël BARBARA : neuroscientifique, directeur de recherche CNRS
Médias
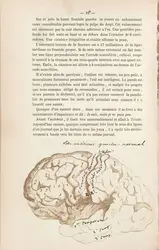
Le centre du langage en 1864
Wellcome Library, Londres ; CC-BY 4.0
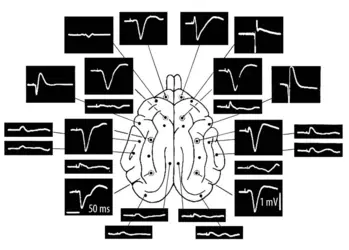
Carte de l’activité cérébrale par électroencéphalographie
1958 Published by Elsevier Ireland Ltd. Reprinted with permission from Elsevier
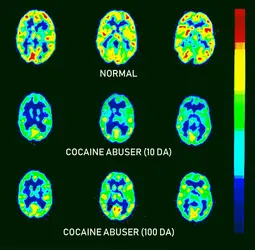
Activité cérébrale estimée par tomographie par émission de positons (TEP)
Brookhaven National Laboratory/ Science Source/ Getty Images
